
UNEP/CMS/COP14/Doc.30.4.4/Annex 3
1
ANNEX 3
CMS Light Pollution Guidelines
Introduction
Natural darkness has a conservation value in the same way that clean water, air and soil has
intrinsic value. Artificial light at night is increasing globally by about two per cent per year (Kyba
et al., 2017). Over the 25-year period 1992 2017 artificial light emissions increased by at
least 49% (Sánchez de Miguel et al., 2021). Animals perceive light differently from humans
and artificial light can disrupt critical behaviour and cause physiological changes in wildlife
(Russart and Nelson, 2018; Sanders et al., 2021). For example, hatchling marine turtles may
not be able to find the ocean when beaches are lit, and fledgling seabirds may not take their
first flight if their nesting habitat never becomes dark (Witherington and Martin, 2003;
Rodríguez et al., 2017c). Tammar wallabies (Macropus eugenii) exposed to artificial light have
been shown to delay reproduction
and clownfish (Amphiprion ocellaris) eggs incubated under
constant light do not hatch (Robert et al., 2015; Fobert et al., 2019).
Consequently, artificial light has the potential to stall the recovery of a threatened species. For
long-distance migrations integral to its life cycle.
Artificial light at night provides for human safety, amenity and increased productivity. These
Guidelines do not infringe on human safety obligations. Where there are competing objectives
for lighting, creative solutions may be needed that meet both human safety requirements for
artificial light and threatened and migratory species conservation.
The Guidelines outline the process to be followed where there is the potential for artificial
lighting to affect wildlife. They apply to new projects, lighting upgrades (retrofitting) and where
there is evidence of wildlife being affected by existing artificial light.
The technology around lighting hardware, design and control is changing rapidly and biological
responses to artificial light vary by species, location and environmental conditions. These
Guidelines do not set prescriptive limits on lighting but give best practice recommendations
for lighting design and, broadly take an outcomes approach to assessing and mitigating the
effect of artificial light on wildlife.
Figure 1 Pink anemone fish and marine turtle laying eggs. Photos: Nigel Marsh and Robert
Thorn.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
2
Development of these Guidelines
These Guidelines constitute an adaptation to an international context of the ‘National Light
Pollution Guidelines for Wildlife Including Marine Turtles, Seabirds and Migratory Shorebirds’
developed by the Government of Australia in 2020. Those guidelines were endorsed by the
CMS Conference of the Parties at its 13
th
Meeting (COP13, Gandhinagar, February 2020)
through Resolution 13.5 Light Pollution Guidelines for Wildlife as an aid to CMS Parties for
assessing and managing the impact of artificial light on susceptible wildlife in their jurisdiction.
Through Decision 13.138 Light Pollution Guidelines for Wildlife CMS COP13 also requested
the CMS Secretariat to prepare additional guidelines for adoption by COP14 on how to avoid
and mitigate the indirect and direct negative effects of light pollution for taxa not yet in the
focus of the Guidelines developed by the Government of Australia. In application of this
Decision, the CMS Secretariat, in consultation with the CMS Scientific Council, developed
additional guidelines addressing impacts of light pollution on migratory landbirds and bats for
consideration by COP14.
In the process of integrating the newly developed guidelines with those already endorsed by
COP13, in agreement with the Government of Australia, it was decided to also undertake a
review of the existing guidelines to better adapt them to the international context provided by
CMS, while limiting technical revision to a minimum. While many of the examples and case
studies provided still concern species and situations encountered in Australia, they should be
referred to comparable situations found elsewhere. The present Guidelines are the result of
this integration and adaptation exercise.
How to use these Guidelines
These Guidelines provide users with the theoretical, technical and practical information
required to assess if artificial lighting is likely to affect wildlife and the management tools to
minimise and mitigate that effect. These techniques can be applied regardless of scale, from
small, domestic projects to large-scale industrial developments.
The aim of the Guidelines is that artificial light will be managed so wildlife is:
1. Not disrupted within, nor displaced from, important habitat; and
2. Able to undertake critical behaviours such as foraging, reproduction
migration and dispersal.
The Guidelines recommend:
1. Always using Best Practice Lighting Design to reduce light pollution and minimise the
effect on wildlife.
2. Undertaking an Environmental Impact Assessment for effects of artificial light on
species for which artificial light has been demonstrated to affect behaviour,
survivorship or reproduction.
Technical Appendices
The Guidelines are supported by a series of technical appendices that provide additional
information about Best Practice Lighting Design, What is Light and How Wildlife Perceives it,
Measuring Biologically Relevant Light, and Artificial Light Auditing. There is also a checklist

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
3
for artificial light management, and species-specific information for the management of
artificial light for Marine Turtles, Seabirds, Migratory Shorebirds, Migratory Landbirds and
Bats. The range of species covered in taxa-specific appendices may be broadened in the
future.
Regulatory Considerations for the Management of Artificial
Light around Wildlife
These Light Pollution Guidelines should be followed to ensure all lighting objectives are
adequately addressed. This may require solutions to be developed, applied and tested to
ensure lighting management meets the needs of human safety and wildlife conservation. The
application of the guidelines should be considered in the context of any relevant Standards
frameworks (e.g. Commission CIE) and the regulatory framework
specific to each national, regional or local context. The Case Studies illustrate examples of
how a liquefied natural gas processing plant, a transport authority, a marine research vessel
and a cosmopolitan city have addressed this challenge.
Associated guidance
These Guidelines should be read in conjunction with:
• relevant national legislation
• relevant conservation advice for migratory species and other wildlife
• other relevant environmental legislation, regulations, and policy and guidance
documents
• CIE 150: 2017 Guide on the Limitation of the Effects of Obtrusive Light from Outdoor
Lighting Installations, 2
nd
edition
• Joint IDA-
• IDA Five Principles for Responsible Outdoor Lighting
• Recommendations to keep dark and quiet skies for science and society
produced by the Committee on the Peaceful Uses of Outer Space with particular
-
recommendations to mitigate the impacts of ALAN on humans, flora and fauna
• Dark and Quiet Skies II for Science and Society Working Group Reports
• The Responsible Outdoor Lighting at Night (ROLAN) Manifesto for lighting
professionals
• Declaration in Defence of the Night Sky and the Right to Starlight
• up-to-date scientific literature
• local and Indigenous knowledge.
Wildlife and Artificial Light
Vision is a critical cue for wildlife to orient themselves in their environment, find food, avoid
predation and communicate (Rich and Longcore, 2006). Wildlife also uses the rhythmic
change in natural light non-visually, especially for biological timekeeping (Foster and
Kreitzman, 2005; Kreitzman and Foster, 2010). An important consideration in the
management of artificial light for wildlife is an understanding of how light is perceived by
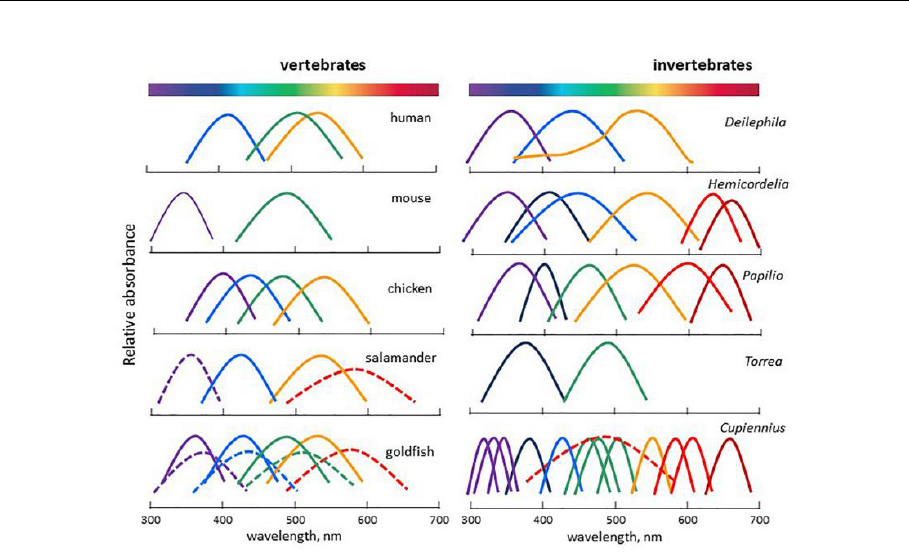
Animals perceive light differently from humans. Most animals are sensitive to ultraviolet
(UV)/violet/blue light, while some birds are sensitive to longer wavelength yellow/orange,
and

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
4
some snakes can detect infra-red (IR) wavelengths
(Figure 2) (Newman and Hartline 1981;
Reed, 1986; Campos, 2017). Understanding the sensitivity of wildlife to different light
wavelengths is critical to assessing the potential effects of artificial light on wildlife.
The way light is described and measured has traditionally focused on human vision. To
manage light appropriately for wildlife, it is critical to understand how light is defined, described
and measured and to consider light from the perspective of the animals concerned.
For a detailed explanation of these issues see What is Light and how does Wildlife Perceive
it? The Glossary provides a summary of terms used to describe light and light measurements
and notes the appropriate terms for discussing the effects of light on wildlife.
Figure 2 Ability to perceive different wavelengths of light in humans and examples of wildlife
taxa are shown by horizontal lines. Black dots represent reported peak sensitivities. Black
dots for bats represent peak sensitivities in an omnivorous bat, based on Winter et al. (2003);
grey dots represent potential peak sensitivities in bats, derived from Feller et al. (2009) and
Simões et al. (2018). Figure adapted from Campos (2017).
How light affects wildlife
Artificial light is known to adversely affect many species
and ecological communities (Bennie
et al., 2016; Gaston et al., 2018; Russart and Nelson, 2018; Sanders and Gaston, 2018). It
can change behaviour and/or physiology, reducing survivorship or reproductive output. It can
also have the indirect effect of changing the availability of habitat or food resources. It can
attract predators and invasive pests, both of which may pose a threat to species of
conservation concern.
Behavioural changes in wildlife have been well described for some species. Adult marine
turtles may avoid nesting on beaches that are brightly lit, and adult and hatchling turtles can
be disoriented and unable to find the ocean in the presence of direct light or skyglow
(Witherington, 1992; Witherington and Martin 2003; Thums et al., 2016; Price et al., 2018).
Similarly, lights can disorient flying birds, particularly during migration, and cause them to

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
5
divert from efficient migratory routes or to collide with infrastructure (Cabrera-Cruz et al.,
2018). Birds may starve when artificial lighting disrupts foraging, and fledgling seabirds may
not be able to take their first flight if their nesting habitat never becomes dark (Rodríguez et
al., 2017c). Migratory shorebirds may use less preferable roosting sites to avoid lights and
may be exposed to increased predation where lighting makes them visible at night (Rodríguez
et al., 2017c).
The stress hormone corticosterone in free living songbirds has been shown to increase when
exposed to white light compared with green or red light and those with high stress hormone
levels have fewer offspring (Ouyang et al., 2015). Plant physiology can also be affected by
artificial light with changes to growth, timing of flowering and resource allocation (Bennie et
al., 2016). This can then have flow-on effects for pollinators and herbivores.
The indirect effects of artificial light can also be detrimental to threatened species. The
Mountain Pygmy Possum (Burramys parvus), for example, feeds primarily on the Bogong
Moth (Agrotis infusa), a long-distance nocturnal migrator that is attracted to light (Warrant et
al., 2016). Recent declines in moth populations, in part due to artificial light, have reduced the
food supply for the possum (Commonwealth of Australia, 2016). Changes in food availability
due to artificial light affect other animals, such as bats (Haddock et al., 2019), and cause
changes in fish assemblages (Bolton et al., 2017). Lighting may also attract invasive pests
such as cane toads (Rhinella marina), or other predators, increasing pressure on species of
conservation concern (González-Bernal et al., 2014; Wilson et al., 2019).
The way in which light affects a species must be considered when developing management
strategies as this will vary on a case-by-case basis.
These Guidelines provide information on the management of artificial light for Marine Turtles,
Seabirds, Migratory Shorebirds, Migratory Landbirds and Bats in the technical appendices.
Consideration should be given to the direct and indirect effect of artificial light on all species
for which artificial light has been demonstrated to negatively affect behaviour, survivorship or
reproduction. If wildlife is present for which there are no demonstrated negative impacts, a
precautionary approach could still be applied as reported patterns could be examples of a
more widespread problem (Davies and Smyth, 2017).
Light Emitting Diodes (LEDs)
During the life of these Guidelines, it is anticipated that light technology may change
dramatically. At the time of writing, LEDs were rapidly becoming the most common light type
used globally. This is primarily because they are more energy efficient than earlier light
sources. LEDs and smart control technologies (such as motion sensors and timers) provide
the ability to control and manage the physical parameters of lighting, making them an integral
tool in managing the effects of artificial light on wildlife.
Whilst LEDs are part of the solution, consideration should be given to some of the
characteristics of LEDs that may influence the effect of artificial light on wildlife. White LEDs
generally contain short wavelength blue light. Short wavelength light scatters more readily
than long wavelength light, contributing more to skyglow. Also, most wildlife is sensitive to blue
light (Figure 2). More detailed consideration of LEDs, their benefits and challenges for use
around wildlife are provided in the Technical Appendix What is Light and how does Wildlife
Perceive it?

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
6
When to Consider the Impact of Artificial Light on Wildlife?
Is Artificial Light Visible Outside?
Any action or activity that includes externally visible artificial lighting should consider the
potential effects on wildlife (refer to Figure 3 below). These Guidelines should be applied at all
stages of management, from the development of planning schemes to the design, approval
and execution of individual developments or activities, through to retrofitting of light fixtures
and management of existing light pollution. Best Practice Lighting Design is recommended as
a minimum whenever artificial lighting is externally visible.
Figure 3 Decision tree to determine whether to undertake an environmental impact
assessment for the effects of artificial light on wildlife.
Best practice lighting design
Natural darkness has a conservation value and should be protected through good quality
lighting design and management for the benefit of all living things. To that end, all infrastructure
that has outdoor artificial lighting or internal lighting that is externally visible should incorporate
best practice lighting design.
Incorporating best practice lighting design into all infrastructure will not only have benefits for
wildlife but will also save energy and provide an economic benefit for light owners and
managers.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
7
Best practice lighting design incorporates the following design principles.
1. Start with natural darkness and only add light for specific purposes.
2. Use adaptive light controls to manage light timing, intensity and colour.
3. Light only the object or area intended – keep lights close to the ground,
directed and shielded to avoid light spill.
4. Use the lowest intensity lighting appropriate for the task.
5. Use non-reflective, dark-coloured surfaces.
6. Use lights without blue, violet and ultraviolet wavelengths if possible. If not,
use lights with reduced or filtered blue, violet and ultraviolet wavelengths.
Figure 4 provides an illustration of best practice lighting design principles. For a detailed
explanation see Technical Appendix Best Practice Lighting Design.
Figure 4 Principles for best practice lighting design.
Is there Important Habitat for Wildlife Located within 20km?
Important habitats are those areas necessary for an ecologically significant proportion of a
species to undertake important activities such as foraging, breeding, roosting or dispersal.
This might include areas that are of critical importance for a particular life stage, are at the
limit of a species range or habitat, or where the species is declining. They may also include
habitat where the presence of light pollution may cause a significant decline in a protected,
threatened or migratory species. Important habitat will vary depending on the species. Special
consideration should be given to the ecological characteristics and values of sites conserved
under international schemes such as the Ramsar and World Heritage conventions,

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
8
International Dark-sky Association Dark Sky Places as well as national and regional schemes
Habitats Directive.
Species specific descriptions of important habitat can be found in Technical Appendices
relating to Marine Turtles, Seabirds, Migratory Shorebirds, Migratory Landbirds and Bats. For
other species see relevant information available in Associated guidance and Desktop Study
of Wildlife.
Where there is important habitat for species that are known to be affected by artificial light
within 20 km of a project, species specific impacts should be considered through an
Environmental Impact Assessment (EIA) process.
The 20 km threshold provides a precautionary limit based on observed effects of skyglow on
marine turtle hatchlings demonstrated to occur at 15-18 km (Hodge et al., 2007; Kamrowski
et al., 2014)
and fledgling seabirds grounded in response to artificial light 15 km away
(Rodríguez et al., 2014). The effect of skyglow may occur at distances greater than 20 km for
some species and under certain environmental conditions. The 20 km threshold provides a
nominal distance at which artificial light impacts should be considered, not necessarily the
distance at which mitigation will be necessary. For example, where a mountain range is
present between the light source and an important turtle nesting beach, further light mitigation
is unlikely to be needed. However, where island infrastructure is directly visible on an important
turtle nesting beach across 25 km of ocean in a remote location, additional light mitigation may
be necessary.
Managing existing light pollution
The impact of artificial light on wildlife will often be the result of the effect of all light sources in
the region combined. As the number and intensity of artificial lights in an area increases there
will be a visible, cumulative increase in skyglow. Skyglow is the brightness of the night sky
caused by the reflected light scattered from particles in the atmosphere. Skyglow comprises
both natural and artificial skyglow. As skyglow increases so does the potential for adverse
impacts on wildlife.
Generally, there is no one source of skyglow and management should be undertaken on a
regional, collaborative basis. Artificial light mitigation and minimisation will need to be
addressed by the community, regulators, councils and industry to prevent the escalation of,
and, where necessary, reduce, the effects of artificial light on wildlife. Light pollution is typically
addressed at the fixture level but should also be managed at the regional level so that lighting
policies and planning are established which ensure the protection of dark areas (See the
recommendations in Part 2. Artificial Light at Night Working Group in UNOOSA, 2021).
Similar to skyglow, local sources of direct light can also affect wildlife, e.g. some insects and
birds are positively phototactic and attracted to artificial lights, while others are negatively
phototactic and avoid ALAN (Van Doren et al., 2017; Owens et al., 2020).
The effect of existing artificial light on wildlife may be identified by protected species managers
or researchers that observe changes in behaviour or population demographic parameters that
can be attributed to increased artificial skyglow and/or direct light. Where this occurs, the
population/behavioural change should be monitored, documented and, where possible, the
source(s) of light identified. An Artificial Lighting Management Plan should be developed in
collaboration with all light owners and managers to mitigate impacts.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
9
Environmental Impact Assessment for Effects of Artificial
Light on Wildlife
There are five steps involved in assessing the potential effects of artificial light on wildlife, and
the adaptive management of artificial light requires a continuing improvement process (Figure
5). The amount of detail included in each step depends on the scale of the proposed activity
and the susceptibility of wildlife to artificial light. The first three steps of the EIA process should
be undertaken as early as possible in the
to inform the project design phase.
Technical Appendices relating to Marine Turtles, Seabirds, Migratory Shorebirds, Migratory
Landbirds and Bats give specific consideration to each of these taxa. However, the process
should also be adopted for other species of conservation concern affected by artificial light.
Qualified personnel
Lighting design/management and the EIA process should be undertaken by appropriately
qualified personnel. Management plans should be developed and reviewed by appropriately
qualified lighting practitioners in consultation with appropriately qualified wildlife biologists or
ecologists.
Figure 5 Flow chart describing the environmental impact assessment process.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
10
Step 1: Describe the project lighting
Describe the existing light environment and characterise the light likely to be emitted from the
site. Information should be collated, including (but not limited to): the location and size of the
project footprint; the number and type of lights; their height, orientation and hours of operation;
site topography and proximity to wildlife and/or wildlife habitat. This information should include
whether lighting will be directly visible to wildlife or contribute to skyglow; the distance over
which this artificial light is likely to be perceptible; shielding or light controls used to minimise
lighting; and spectral characteristics (wavelength) and intensity of lights.
Project specific lighting should be considered in the context of the existing light environment
and the potential for cumulative effects of multiple light sources. The information collected
should be sufficient to assess the likely effects of artificial light on wildlife given the biology
and ecology of species present (Step 2).
Where there will be a need to monitor the effectiveness of artificial light mitigation and
management strategies (Step 5), baseline monitoring will be necessary. Measurements of the
existing light environment should recognise and account for the biologically relevant short
(violet/blue) and long (orange/red) wavelengths of artificial lighting (see Measuring Biologically
Relevant Light).
Lighting objectives
During the planning phase of a project the purpose of artificial lighting should be clearly
articulated, and consideration should be given as to whether artificial light is required at all.
Lighting objectives should be specific in terms of location and times for which artificial light is
necessary, whether colour differentiation is required and whether some areas should remain
dark. The objectives should include the wildlife requirements identified in Step 2 and be
consistent with the aims of these Guidelines.
For more information about developing lighting objectives see Best Practice Lighting Design.
Step 2: Describe wildlife
Describe the biology and ecology of wildlife in the area that may be affected by artificial light
(species identified during the screening process, Figure 3). The abundance, conservation
status and regional significance of wildlife will be described, as will the location of important
habitat. Recognise biological and ecological parameters relevant to the assessment,
physiological sensitivity to wavelength and intensity, and its visual field.
Depending on the availability of information, scale of the activity and the susceptibility of
wildlife to artificial light, this step may only require a desktop analysis. Where there is a paucity
of information or the potential for effects is high, field surveys may be necessary. Where there
will be a need to monitor the effectiveness of lighting mitigation and management strategies
(Step 5), baseline monitoring will be necessary.
Desktop study of wildlife
A review of the available government databases, scientific literature and unpublished reports
should be conducted to determine whether wildlife of conservation concern that is susceptible
to the effects of artificial light and/or important habitat could be present within 20 km of the
area of interest.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
11
To assess the risks to a species, an understanding of its susceptibility to the effects of light
should be evaluated, as well as the potential for artificial light to affect the local population.
The species conservation status should be identified. Relevant population demographic and
behavioural characteristics that should be considered include population size, life stages
present and normal behaviour in the absence of artificial light. This step should also identify
biological and ecological characteristics of the species that will be relevant to the assessment.
This may include understanding the seasonality of wildlife using the area; behaviour (i.e.
reproduction, foraging, resting, nocturnality); migratory pathways; and life stages most
susceptible to artificial light. Consideration should also be given to how artificial light may affect
food sources, availability of habitat, competitors or predators.
Field surveys for wildlife
Where there are insufficient data available to understand the actual or potential importance of
a population or habitat it may be necessary to conduct field surveys. The zone of influence for
artificial lighting will be case and species specific. Surveys should describe habitat, species
abundance and density on a local and regional scale at a biologically relevant time of year.
As well as field surveys, remote sensing methods can be applied. For some taxa, e.g. birds,
information from weather radar systems, or even dedicated bird radar, will greatly increase
the robustness and scope of surveys, especially during the night. Such data is invaluable for
understanding broadscale patterns of movements, especially relative to light pollution.
Baseline monitoring
Where it is considered likely that artificial lighting will impact wildlife, it may be necessary to
undertake baseline monitoring to inform mitigation and light management (Step 5).
Field survey techniques and baseline monitoring needs will be species specific and detailed
parameters and approaches are described in the Marine Turtles, Seabirds, Migratory
Shorebirds, Migratory Landbirds and Bats Technical Appendices. Guidance from species
experts should be sought for other species.
Step 3: Risk assessment
Using information collated in steps one and two, the level of risk to wildlife should be assessed.
Risk assessments should be undertaken on a case-by-case basis as they will be specific to
the wildlife involved, the lighting objectives and design, and the prevailing environmental
conditions. Assessments should be undertaken in accordance with relevant risk management
guidelines. The scale of the assessment is expected to be commensurate with the scale of
the activity and the vulnerability of the wildlife present.
In general, the assessment should consider how important the habitat is to the species (e.g.
is this the only place the animals are found), the biology and ecology of wildlife, the amount
and type of artificial light at each phase of development (e.g. construction/operation) and
whether the lighting scenario is likely to cause an adverse response. The assessment should
take into account the artificial light impact mitigation and management that will be
implemented. It should also consider factors likely to
distance to the lighting source; and whether light will be directly visible or viewed as skyglow.
The process should assess whether wildlife will be disrupted or displaced from important
habitat, and whether wildlife will be able to undertake critical behaviours such as foraging,
reproduction, and dispersal.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
12
Where a likely risk is identified, either the project design should be modified, or further
mitigation put in place to reduce the risk.
If the residual risk is likely to be significant, consideration should be given as to whether the
project should be referred for assessment under relevant national or subnational legislation.
Step 4: Artificial lighting management plan
The management plan will document the EIA process. The plan should include all relevant
information obtained in Steps 1-3. It should describe the lighting objectives; the existing light
environment; susceptible wildlife present, including relevant biological characteristics and
behaviour; and proposed mitigation. The plan should clearly document the risk assessment
process, including the consequences that were considered, the likelihood of occurrence and
any assumptions that underpin the assessment. Where the risk assessment deems it unlikely
that the proposed artificial light will affect wildlife and an artificial lighting management plan is
not required, the information and assumptions underpinning these decisions should be
documented.
Where an artificial lighting management plan is deemed necessary, it should document the
scope of monitoring and auditing to test the efficacy of proposed mitigation and triggers to
revisit the risk assessment. This should include a clear adaptive management framework to
support continuous improvement in light management, including a hierarchy of contingency
management options if biological and light monitoring or compliance audits indicate that
mitigation is not meeting the objectives of the plan.
The detail and extent of the plan should be proportional to the scale of the development and
potential impacts to wildlife.
A toolbox of species-specific options are provided in the Marine Turtles, Seabirds, Migratory
Shorebirds, Migratory Landbirds and Bats Technical Appendices. Guidance from species
experts should be sought for other species.
Step 5: Biological and light monitoring and auditing
The success of the impact mitigation and artificial light management should be confirmed
through monitoring and compliance auditing. Light audits should be regularly undertaken, and
biological and behavioural monitoring should take place on a timescale relevant to the species
present. Observations of wildlife interactions should be documented and accompanied by
relevant information such as weather conditions and moon phase. Consideration should be
given to monitoring control sites. Monitoring should be undertaken both before and after
changes to artificial lighting are made at both the affected site and the control sites. The results
of monitoring and auditing are critical to an adaptive management approach, with the results
used to identify where improvements in lighting management may be necessary. Audits should
be undertaken by appropriately qualified personnel.
Baseline, construction or post construction artificial light monitoring, wildlife biological
monitoring and auditing are detailed in Measuring Biologically Relevant Light, Light Auditing
and species-specific Marine Turtles, Seabirds, Migratory Shorebirds, Migratory Landbirds and
Bats Technical Appendices.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
13
Step 6: Review
Once light audits and biological monitoring have been completed, a review of whether the
lighting objectives have been met should be conducted. The review should incorporate any
changing circumstances and make recommendations for continual improvement. The
recommendations should be incorporated through upgraded mitigations, changes to
procedures and renewal of the light management plan.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
14
Case Studies
Unlike many forms of pollution, artificial light can be removed from the environment. The
following case studies show it is possible to balance the requirements of both human safety
and wildlife conservation.
Gorgon Liquefied Natural Gas Plant on Barrow Island, Western Australia
The Chevron-atural gas projects. The
liquefied natural gas (LNG) processing facility is on Barrow Island a Western Australian Class
A nature reserve off the Pilbara Coast known for its diversity of fauna, including important
nesting habitat for flatback turtles (Natator depressus) (Moro et al., 2018).
The LNG plant was built adjacent to important turtle nesting beaches. The effect of light on
the turtles and emerging hatchlings was considered from early in the design phase of the
project and species-specific mitigation was incorporated into project planning (Moro et al.,
2018). Light management is implemented, monitored and audited through a light management
plan and turtle population demographics and behaviour through the Long Term Marine Turtle
Management Plan (Chevron Australia, 2018).
Lighting is required to reduce safety risks to personnel and to maintain a safe place of work
under workplace health and safety requirements. The lighting objectives considered these
requirements while also aiming to minimise skyglow and eliminate direct light spill on nesting
beaches. This includes directional or shielded lighting, the mounting of light fittings as low as
practicable, louvered lighting on low level bollards, automatic timers or photovoltaic switches
and black-out blinds on windows. Accommodation buildings were oriented so that a minimal
number of windows faced the beaches and parking areas were located to reduce vehicle
headlight spill onto the dunes.
Lighting management along the LNG jetty and causeway adopted many of the design features
used for the plant and accommodation areas. LNG loading activity is supported by a fleet of
tugs that were custom built to minimise external light spill. LNG vessels are requested to
minimise non-essential lighting while moored at the loading jetty.
To reduce skyglow, the flare for the LNG plant was designed as a ground box flare, rather
than the more conventional stack flare. A louvered shielding wall further reduced the effects
of the flare.
Lighting reviews are conducted prior to the nesting season to allow time to implement
corrective actions if needed. Workforce awareness is conducted at the start of each turtle
breeding season to further engage the workforce in the effort to reduce light wherever
possible.
The Long Term Marine Turtle Management Plan
provides for the ongoing risk assessment of
the impact of artificial light on the flatback turtles nesting on beaches adjacent to the LNG
plant, including mitigation measures to minimise the risk from light to turtles (Chevron
Australia, 2018). The plan also provides for an ongoing turtle research and monitoring
programme. The plan is publicly available.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
15
Figure 6 Liquefied natural gas plant on Barrow Island. Photo: Chevron Australia.
Phillip Island, Victoria, Australia
short-tailed
shearwaters (Ardenna tenuirostris). It supports more than six per cent of the global population
of this species (Rodríguez et al., 2014). Shearwaters nest in burrows and are nocturnally
active at their breeding colonies. Fledglings leave their nests at night. When exposed to
artificial light fledglings can be disoriented and grounded. Some fledglings may reach the
ocean, but then be attracted back toward coastal lighting. Fledglings are also vulnerable to
collision with infrastructure when disoriented and once grounded become vulnerable to
predation or roadkill
(Figure 7) (Rodríguez et al., 2017c).
Phillip Island also attracts over a million visitors a year during peak holiday seasons to visit
the Little Penguin (Eudyptula minor) ecotourism centre, the Penguin Parade
®
. Most visitors
drive from Melbourne across a bridge to access the island. The increase in road traffic at
sunset during the Easter break coincides with the maiden flight of fledgling shearwaters from
their burrows (Rodríguez et al., 2014).
In response to the deaths of fledglings, Phillip Island Nature Parks has an annual shearwater
rescue programme to remove and safely release grounded birds (Rodríguez et al., 2014). In
collaboration with SP Ausnet and Regional Roads Victoria, road lights on the bridge to the
island are turned off during the fledgling period (Rodríguez et al., 2017b). To address human
safety concerns, speed limits are reduced and warning signals put in place during fledgling
season (Rodríguez et al., 2017ab). The reduced road lighting and associated traffic controls
and warning signals, combined with a strong rescue programme, have reduced the mortality
rate of shearwaters (Rodríguez et al., 2014).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
16
Figure 7 Short-Tailed Shearwater (Ardenna tenuirostris) fledgling grounded by artificial light,
Phillip Island. Photo: Airam Rodríguez.
Raine Island research vessel light controls, Queensland, Australia
The Queensland Marine Parks primary vessel Reef Ranger is a 24 m catamaran jointly funded
by the Great Barrier Reef Marine Park Authority and the Queensland Parks and Wildlife
Service under the Field Management Program (FMP). The Reef Ranger is often anchored at
offshore islands that are known marine turtle nesting sites and is regularly at Raine Island,
turtle (Chelonia mydas) nesting sites (Limpus et al., 2003)
and
a significant seabird rookery.
Vessels often emit a lot of artificial light when at anchor and the FMP took measures to
minimise direct lighting spillage from the vessel. A lights-off policy around turtle nesting
beaches was implemented, where the use of outdoor vessel lights was limited, except for
safety reasons.
The original fit out of the vessel did not include internal block-out blinds (Figure 8A). These
were installed before the 2018-19 Queensland turtle nesting season. The blinds stop light
being emitted from inside the vessel, therefore limiting light spill around the vessel (Figure 8B).
This can make an important difference at remote (naturally dark) sites such as Raine Island.
Anecdotal evidence suggests hatchlings previously attracted to, and captured in, light pools
around the vessel are no longer drawn to the Reef Ranger.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
17
Figure 8 Vessel lighting management at Raine Island A. Vessel with decking lights, venetian
blinds down and anchor light on; and B. Vessel with outside lights off, and block-out blinds
installed (note the white anchor light is a maritime safety requirement). Photo: Queensland
Parks and Wildlife Service.
“Tribute in Light”, New York, USA
The Tribute in Light TiL) is an event held annually since 2002 on September 11th to
remember the lives lost during the terrorist attacks of September 11th, 2001. The National
September 11 Memorial & Museum (NSMM) currently operates the light installation on top of
a parking garage near the site of the former World Trade Center in New York City (NYC), NY
at the southern end of Manhattan Island. NYC is a heavily light polluted environment, but even
in this location, 88 ~7,500 watt Xenon bulbs pointing skyward to zenith are visible for at least
100 km on a clear night, giving the appearance of two tall towers of light. The bulbs have a
dichroic treatment as well as nickel rhodium reflectors that significantly reduce infrared and
ultraviolet spectra and create an effect similar to daylight.
Massive nocturnal migratory movements of birds regularly occur over the area during mid-
September (see BirdCast; Farnsworth et al., 2016; van Doren et al., 2015; Horton et al.,
2016ab). induced significant behavioural
alterations in birds, even in good visibility (i.e., clear skies without cloud cover) conditionsto
altitudes up to 4 km
As the timing of migratory movements depends on local and regional weather and wind
conditions (Richardson, 1978; Van Belle et al., 2007; Kemp et al., 2013; La Sorte et al., 2015),
the magnitude of migratory passage on the single night of September 11th varies greatly
across years. An existing agreement between New York City Audubon (NYCA) and NSMM
governs when to initiate the shutdown procedures: when numbers of birds circling in the
beams exceed 1,000 individuals, based on visual observations, NYCA requests that the TiL
y
monitoring birds and their behaviours in the beams. This plan was implemented before any
data supported its efficacy.
The study by Van Doren et al. (2017) quantified:
1) densities and flight speeds of aerial migrants near the light installation using weather
surveillance radar data, revealing how numbers of birds and their rates of passage
changed in the presence or absence of illumination,
2) -flight vocalizations, or flight calls, from the
base of the installation, and

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
18
3) simulated bird behaviours modelled in ALAN conditions for comparison with observed
radar data.
The simple conclusion was that high intensity lights have the ability to greatly impact avian
migratory behaviour under a wide range of conditions. Van Doren et al. (2017) observed that
when the installation was illuminated, birds aggregated in high densities, decreased their flight
speeds, followed circular flight paths, and vocalised frequently when the installation was
period of 7 days over 7 years. Bird densities near the TiL installation exceeded magnitudes
20-
However, behavioural disruptions disappeared when lights were extinguished, highlighting
that removal of light during nights with substantial bird migration is a viable strategy for
minimising potentially fatal interactions between ALAN, structures, and birds.
most recognisable features of the nocturnal lightscape of NYC, a lightscape with enormous
public recognition globally. Yet, a hallmark of this study was frequent and public cooperation
among the NSMM, the Municipal Arts Society, NYCA, the Cornell Lab of Ornithology, and
stakeholders with direct interest and responsibility for this event, all of whom acknowledged
its potential to negatively impact birds, to shut off the lights periodically for the benefit of
migratory birds. This is an encouraging acknowledgment of the importance of bird
conservation. Mo
viewers, media coverage often highlighted a unified message from stakeholders about
balancing potential hazards to migrating birds with the intent and spirit of the display. All parties
agreed to keep the display illuminated unless potentially hazardous conditions for birds
necessitated a short-term shutdown of the lights. Whereas discontinuing the display would be
best for nocturnally migrating birds, such a scenario may never be possible given the
psychological and social needs of the local, regional, national and global human communities.
There has been significant positive media coverage of this study related to the TiL, including
among scientists, print media, cinematic productions, internal and social media, and non-
fiction books, covering the consensus building, the protection of migrating birds, the
methodology, and the impacts for actions to reduce light pollution. Moreover, the profile,
discussion, and attention surrounding the study of its impacts continues to provide 1)
groundwork for mitigating impacts to birds at the location annually and, more importantly, 2)
science applied to support the passage of critical legislation by the New York City Council to
reduce or eliminate light pollution to protect nocturnally migrating birds.
TiL is an outstandingly intense light source, but recent studies both in the Americas and
Europe (Van Doren et al. 2021, Korner et al., 2022) have confirmed the massive scale of threat
for migratory birds in more typical urban settings. Using long-term data from one building each,
both studies provided evidence for high casualties among nocturnally migrating landbirds
through attraction by nocturnal illumination.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
19
Appendix A Best Practice Lighting
Design
Natural darkness has conservation value in the same way as clean water, air and soil
and should be protected through good quality lighting design.
Simple management principles can be used to reduce light pollution, including:
1. Start with natural darkness and only add light for specific purposes.
2. Use adaptive light controls to manage light timing, intensity and colour.
3. Light only the object or area intended – keep lights close to the ground,
directed and shielded to avoid light spill.
4. Use the lowest intensity lighting appropriate for the task.
5. Use non-reflective, dark-coloured surfaces.
6. Use lights without blue, violet and ultraviolet wavelengths if possible. If not,
use lights with reduced or filtered blue, violet and ultraviolet wavelengths.
The application of best practice lighting design for all outdoor lighting is intended to reduce
skyglow and minimise the effects of artificial light on wildlife.
Lighting Objectives
At the outset of a lighting design process, the purpose of artificial lighting should be clearly
stated and consideration should be given as to whether it is required at all. Exterior lighting for
public, commercial or industrial applications is typically designed to provide a safe working
environment. If a safety concern exists, alternatives to outdoor lighting should be used where
possible, for example curbs, steps and other potential hazards can be highlighted using
reflective paints and/or tapes and/or self-luminous materials rather than installing lighting (IDA
and IES, 2020).
Exterior lighting may also be required to provide for human amenity or commerce. Conversely,
areas of darkness, seasonal management of artificial light, or minimised skyglow may be
necessary for wildlife protection, astronomy or dark sky tourism.
Lighting objectives will need to consider the regulatory requirements and standards relevant
to the activity, location and wildlife present.
Objectives should be described in terms of specific locations and times for which artificial light
is necessary. Consideration should be given to whether colour differentiation is required and
if some areas should remain dark either to contrast with lit areas or to avoid light spill. Where
relevant, wildlife requirements should form part of the lighting objectives for example by
avoiding the illumination of vegetation.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
20
Façade lighting (also known as vanity lighting, architectural lighting or decorative lighting)
should not be used or should be eliminated where possible. The lighting of building façades,
for example churches, often contributes to light pollution in the surrounding area and has been
highlighted as affecting roost sites of bats, particularly throughout Europe. See Appendix J -
Bats. The illumination of monuments in rural areas should be avoided in particular. If façade
lighting is to be used the light should be completely confined to the target surface and subject
to illuminance or luminance upper limits (Kyba et al., 2018).
A lighting installation will be deemed a success if it meets the lighting objectives (including
wildlife needs) and areas of interest can be seen by humans clearly, easily, safely and without
discomfort.
The following provides general principles for lighting that will benefit the environment, local
wildlife and reduce energy costs.
Principles of Best Practice Lighting Design
Good lighting design incorporates the following design principles. They are applicable
everywhere, especially in the vicinity of wildlife.
1. Start with natural darkness
The starting point for all lighting designs should be natural darkness (Figure 9). Artificial light
should only be added for specific and defined purposes, and only in the required location and
for the specified duration of human use. Designers should consider an upper limit on the
amount of artificial light and only install the amount needed to meet the lighting objectives.
In a regional planning context, consideration should be given to
where activities that involve outdoor artificial light are prohibited under local planning schemes.
Figure 9 Start with natural darkness.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
21
2. Use adaptive controls
Recent advances in smart control technology provide a range of options for better controlled
and targeted artificial light management (Figure 10). For example, traditional industrial lighting
may need to remain illuminated all night because High-Pressure Sodium, metal halide, and
fluorescent lights have a long warm up and cool down period which could jeopardise operator
safety in the event of an emergency. With the introduction of smart controlled LED lights, plant
lighting can be switched on and off instantly and activated only when needed, for example,
when an operator is physically present within the site.
Smart controls and LED technology allow for:
•remotely managing lights (computer controls)
•instant on and off switching of lights
•control of light colour (emerging technology)
•dimming, timers, flashing rate, motion sensors, well defined directivity of light.
Adaptive controls should maximise the use of the latest lighting technology to minimise
unnecessary light output and energy consumption. Controls should be automatic with failsafe
switches which do not require a human to switch them off every night. There should be no
capacity for such lighting to be accidentally left on all night. Businesses and offices should use
adaptive controls to turn off lights after usual business hours and to limit illuminated signage
brightness and surface area.
Streetlights can use adaptive lighting control (dimming, detection, scheduling) to dim the
lighting levels according to the relevant lighting classes (for example see CIE 115:2010:
Lighting of Roads for Motor and Pedestrian Traffic).
Figure 10 Use adaptive controls to manage light timing, intensity and colour.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
22
3. Light only the intended object or area - keep lights close to the ground,
directed and shielded
Light spill is light that falls outside the area intended to be lit. Light that spills above the
horizontal plane contributes directly to artificial skyglow while light that spills into adjacent
areas on the ground (also known as light trespass) can be disruptive to wildlife in adjacent
areas. All light fittings should be located, directed or shielded to avoid lighting anything but the
target object or area (Figure 11). Existing lights can be modified by installing a shield.
Figure 11 Lights should be shielded to avoid lighting anything but the target area or object.
Figure adapted from Witherington and Martin (2003).
Lower height lighting that is directional and shielded can be extremely effective. Light fixtures
should be located as close to the ground as possible and shielded to reduce skyglow (Figure
12).
Figure 12 Walkway lighting should be mounted as low as possible and shielded. Figure
adapted from Witherington and Martin (2003).
Artificial light can be prevented from shining above the horizontal plane by ensuring the
luminaire is mounted horizontally relative to the ground and not at an angle, or mounted on a
building so that the structure prevents the light shining above the horizontal plane, for example
recess a light into an overhanging roof eave. When determining angle of the mounting,
consideration should be given to the reflective properties of the receiving environment. The
upward light output ratio (ULOR) should be as close to 0.0% as possible (See CIE 150:2017
Guide on the Limitation of the Effects of Obtrusive Light from Outdoor Lighting Installations).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
23
This requires luminaires to be mounted horizontally and have flat optics below the light source
(COPUOS, 2021). Lighting with adjustable mounts should not be used as that allows
luminaires to be tilted upwards, thereby defeating their proper shielding.
For streetlights, efficient lighting design including the proper selection of optics and luminous
power should be used, avoiding shining direct light onto roadway and sidewalk surfaces.
Shielding should be used where necessary and according to traffic volume and conditions.
Lighting pole distance and height should be selected via proper lighting design, in a way that
minimises spill light, glare and the illumination of the surrounding area while respecting the
relevant illumination limits.
If an unshielded fitting is to be used, consideration should be given to the direction of the light
and the need for some form of permanent physical opaque barrier that will provide the
shielding requirement. This can be a cover or part of a building (Figure 13). Care should be
taken to also shield adjacent surfaces, if they are lightly coloured, to prevent excessive
reflected light from adding to skyglow.
Consideration should also be given to blocking light spill from internal light sources. This
should include block-out blinds, curtains or shutters for transparent portions of a building,
including sky lights. Some locations and climates may not allow for this due to lack of available
technology and other practicalities, for example in places where air conditioning is not
available it may be necessary to open windows for airflow which means that blocking light spill
from internal sources may not be possible.
Floodlighting should be avoided as much as possible. When it is used it should be top-down
and fully shielded. See the IDA-Criteria for Community-Friendly Outdoor Sports Lighting for
further advice.
Searchlights should only be used for emergency situations.
Brightness of LED signs and digital billboards should be limited. For best management
practices regarding LED signs see the IDA Guidance for Electronic Message Centers (EMCs)
(IDA, 2019) and Zielinska-Dabkowska and Xavia (2019).
Figure 13 Lighting should be directed to ensure only the intended area is lit. Figure adapted
from Witherington and Martin (2003).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
24
4. Use appropriate lighting
Lighting intensity should be appropriate for the activity. Starting from a base of no lights, use
only the minimum number and intensity of lights needed to provide safe and secure
illumination for the area at the time required to meet the lighting objectives. The minimum
amount of light needed to illuminate an object or area should be assessed during the early
design stages and only that amount of light installed. For example, Figure 14 provides options
from best to worst for lighting for a parking lot.
Figure 14 Lighting options for a parking area. Figure adapted from Witherington and Martin
(2003).
Off-the-shelf lighting design models
Use of computer design engineering packages that do not include wildlife needs and only
recommend a standard lighting design for general application should be avoided or modified
to suit the specific project objectives, location and risk factors.
Consider the intensity of light produced rather than the energy required to make it
Improvements in technology mean that new bulb types produce a significantly greater amount
of light per unit of energy. For example, LED lights produce between two and five times the
amount of light as incandescent bulbs. The amount of light produced (lumen), rather than the
amount of energy used (watt) is the most important consideration in ensuring that an area is
not over lit.
Consider re-evaluating security systems and using motion sensor lighting
Technological advances mean that techniques such as computer managed infrared tracking
of intruders in security zones is likely to result in better detection rates than a human observer
monitoring an illuminated zone. However, some wildlife is sensitive to infrared (IR) and near-
IR and, therefore, these emissions should be eliminated wherever possible (Campbell et al.,
2002; Shcherbakov et al., 2013; UNOOSA, 2020).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
25
Use low glare lighting
High quality, low glare lighting should always be a strong consideration regardless of how the
project is to be designed. Low glare lighting enhances visibility for the user at night, reduces
eye fatigue, improves night vision and delivers light where it is needed.
Using low mounting heights also works well as a means of reducing or eliminating glare. For
example, lighting a pathway with low, bollard-style lighting that confines light to the path
surface virtually eliminates glare compared to the use of conventional, post-top lighting.
5. Use non-reflective, dark coloured surfaces
Light reflected from highly polished, shiny or light-coloured surfaces such as white painted
infrastructure, polished marble or white sand can contribute to skyglow. For example,
alternatives to painting storage tanks with white paint to reduce internal heating should be
explored during front-end engineering design. In considering surface reflectance, the need to
view the surface should be taken into consideration as darker surfaces will require more light
to be visible. It should also be noted that using reflective surfaces can sometimes reduce the
need for additional lighting. Reflective surfaces should be used or avoided appropriately and
in a manner that reduces overall light pollution. Reflection from other surfaces, like
pavements, can also be minimised by carefully selecting materials. The colour of paint or
material selected should be included in the Artificial Lighting Management Plan. Open water
should not be illuminated because it reflects light directly upward into the night sky and
shorter wavelengths can penetrate into water thereby impacting aquatic wildlife.
Figure 15 Use non-reflective dark coloured surfaces.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
26
6. Use lights without blue, violet and ultraviolet wavelengths if possible. If not,
use lights with reduced or filtered blue, violet and ultraviolet wavelengths.
Short wavelength light (blue) scatters more readily in the atmosphere and therefore
contributes more to skyglow than longer wavelength light. Furthermore, most wildlife is
sensitive to short wavelength (blue/violet) light (for detailed discussion see What is Light and
how does Wildlife Perceive it?). As a general rule, only lights with little or no short wavelength
(500 nm and below) violet or blue light should be used to avoid unintended effects. Where the
wildlife concerned is sensitive to longer wavelength light (e.g. some bird species),
consideration should be given to wavelength selection on a case-by-case basis.
When determining the appropriate wavelength of light to be used, all lighting objectives should
be taken into account. If good colour rendition is required for human use, then other mitigation
measures such as tight control of light spill, use of head torches, or timers or motion sensors
to control lights should be implemented.
It is not possible to tell how much blue light is emitted from an artificial light source by the
colour of light it produces (see Light Emitting Diodes). LEDs of all colours, particularly white,
can emit a high amount of blue light and the Correlated Colour Temperature (CCT) only
provides a proxy for the blue light content of a light source. Consideration should be given to
the spectral characteristics (spectral power distribution) of the light source to ensure short
wavelength (500 nm and below) light is minimised. Longer wavelengths (red) tend to not
scatter as far and may affect a smaller area. However, many species are vulnerable to
exposure to longer wavelengths or infrared radiation (IR). As IR is not visible for humans, it
should not be used in outdoor lighting. Older traditional light sources such as HID, HPS and,
to a certain extent, even fluorescent lamps emit IR. The use of LEDs means IR can be
eliminated from outdoor lighting.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
27
Appendix B What is Light and how
does Wildlife Perceive it?
A basic understanding of how light is defined, described and measured is critical to
designing the best artificial light management for the protection of wildlife.
Humans and animals perceive light differently. However, defining and measuring light
has traditionally focused exclusively on human vision. Commercial light monitoring
equipment is calibrated to the sensitivity of the human eye and has poor sensitivity to
the short wavelengths that are most visible to wildlife. Impacts of artificial light on
wildlife vary by species and should be considered on a case-by-case basis. These
issues should be considered when describing, monitoring and designing lighting near
important wildlife habitat. The higher the intensity of light, the more likely that there will
be ecological impacts, so keeping intensity low is critically important.
What is Light?
Light is a form of energy and is a subset of the electromagnetic spectrum that includes visible
light, microwaves, radio waves and gamma rays (Figure 16). In humans, visible light ranges
from 380 nm to 780 nm - between the violet and red regions of the electromagnetic spectrum.
In animals, visibility ranges from 300 nm to greater than 700 nm, depending on the species.
White light is a mixture of all wavelengths of light ranging from short wavelength blue to long
wavelength red light.
The perception of diff
characterised by how the human eye perceives light, ranging from red (700 nm), orange (630
nm), yellow (600 nm), green (550 nm), blue (470 nm), indigo (425 nm) and violet (400 nm)
(Figure 16). Generally, this is not how animals see light (Figure 2). Importantly, light affects
wildlife not only through visual pathways but also through photoreceptors for example in the
brain or associated glands (Falcón et al., 2020). This non-visual light perception directly acts
on animals via physiological pathways, such as the circadian system and other forms of
biological rhythms.
Figure 16 The electromagnetic spectrum. The 'visible light spectrum' occurs between 380-780
nm and is the part of the spectrum that the human eye can see. Credit: Mihail Pernichev
(Iristech, 2018).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
28
Artificial light
Artificial light at night has many positive attributes. It can enhance human safety and provide
for longer periods of work or recreation. However, it can also have negative effects. For
example, it can cause:
• physiological damage to retinal cells in human and animal eyes (Algvere et al., 2006),
• disruption of the circadian cycles in vegetation, animals and humans (West et al., 2010;
Bennie et al., 2016; Russart and Nelson, 2018),
• changes in animal orientation, feeding or migratory behaviour (Bird et al., 2004;
Salmon, 2006; Pendoley and Kamrowski, 2015a; Warrant et al., 2016).
The biological mechanisms that cause these effects vary. It is necessary to understand some
basic light theory and language in order to assess and manage the effect of light on wildlife.
Some basic principles are briefly described in this section.
Vision in Animals
Vision is a critical cue for animals to orient themselves in their environment, find food, avoid
predation and communicate (Rich and Longcore, 2006). Humans and wildlife perceive light
differently. Some animals do not see long wavelength red light at all, while others see light
beyond the blue-violet end of the spectrum and into the ultraviolet (Figure 17).
Both humans and animals detect light using photoreceptor cells in the eye called cones and
rods. Colour differentiation occurs under bright light conditions (daylight). This is because
bright light activates the cones and it is the cones that allow the eye to see colour. This is
known as photopic vision.
Under low light conditions (dark adapted vision), light is detected by cells in the eye called
rods. Rods only perceive light in shades of grey (no colour). This is known as scotopic vision
and it is more sensitive to shorter wavelengths of light (blue/violet) than photopic vision.
The variation in the number and types of cells in the retina means animals and humans do not
of wavelengths means they can perceive light at that wavelength, and it is likely they will
respond to that light source.
UNEP/CMS/COP14/Doc.30.4.4/Annex 3
29
Figure 17 Ability to perceive different wavelengths of light in humans and animal species.
Figure from Falcón et al. (2020), adapted and modified from Imamoto and Shichida (2014),
Warrant (2019).
Sensitivity to blue light
Sensitivity to high energy, short wavelength UV/violet/blue light is common in wildlife (Figure
17). This light is strongly detected under scotopic (dark adapted) vision, particularly in
nocturnal species. Short wavelength light at the blue end of the spectrum has higher energy
than longer wavelength light at the red end of the spectrum. This is important for understanding
the physical impact that the short wavelength, high energy UV/blue light has on damaging
photoreceptor cells in the human eye (Tosini et al., 2016). Although not well described in
wildlife, it is not unreasonable to expect that at high intensities blue light has the potential to
damage photoreceptors in wildlife.
In addition to the potential for physical damage to the eye from exposure to blue light (400 -
490 nm), there is mounting evidence that exposure to these wavelengths at night may affect
human and wildlife physiological functions. This is because a third type of photoreceptor cell
has recently been identified in the retina of the mammalian eye the photosensitive retinal
ganglion cells (pRGCs). The pRGCs are not involved in image-forming vision (this occurs in
the rods and cones), but instead are involved in the regulation of melatonin and in
synchronising circadian rhythms to the 24-hour light/dark cycle in animals (Ecker et al., 2010).
These cells are particularly sensitive to blue light (Berson, 2007). In non-mammalian
vertebrates, light is also perceived in various parts of the brain and in particular in the pineal
and para-pineal glands, which are the main secretion sites for the hormone melatonin
(Grubisic et al., 2019; Falcón et al., 2020). Melatonin is a hormone found in plants, animals
and microbes. Changes in melatonin production can affect daily behaviours such as bird
waking (de Jong et al., 2015), foraging behaviour and food intake (Angers et al., 2003)
and
seasonal cues such as the timing of reproduction in animals, causing off-spring to be born
during non-optimal environmental conditions (Robert et al., 2015).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
30
Factors Affecting Perception of Light
Factors affecting how wildlife perceives light include the type of cells being employed to detect
light (photopic vs scotopic vision); whether the light is viewed directly from the source or as
reflected light; how the light interacts with the environment; and the distance from the light
source. These influences are discussed below.
Perspective
field. For instance, when flying, birds will generally be looking down on artificial light sources,
stretch around to almost behind their head.
Bright vs dim light
Understanding photopic and scotopic vision is important when selecting the colour
(wavelength) and intensity of a light. In animals scotopic (dark adapted) vision allows for the
detection of light at very low intensities (Figure 18). This dark adaption may explain why
nocturnal wildlife are extremely sensitive to white and blue light even at low intensities.
Direct vs reflected
Understanding the difference between light direct from the source (luminance) and how much
incident light illuminates a surface (illuminance) is important when selecting methods for
measuring and monitoring light. Equipment used to measure illuminance and luminance is not
interchangeable and will lead to erroneous conclusions if used incorrectly.
Luminance describes the light that is emitted, passing through or reflected from a surface that
is detected by the human eye. The total amount of light emitted from a light is called luminous
flux and represents the light emitted in all directions (Figure 19). Luminance is quantified using
a Spectroradiometer or luminance meter.
Illuminance measures how much of the incident light (or luminous intensity) illuminates a
surface. Illuminance is quantified using an Illuminance spectrophotometer or Lux meter.
Figure 18 Scotopic and photopic luminosity functions in humans. Data source: Luminosity
functions.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
31
Figure 19 Luminous flux, luminance and illuminance.
Visibility of light in the environment
The physical properties of light include reflection, refraction, dispersion, diffraction and
scattering. These properties are affected by the atmosphere through which light travels. Short
wavelength violet and blue light scatters in the atmosphere more than longer wavelength light
such as green and red, due to an effect known as Rayleigh scattering (Benenson et al., 2006).
Scattering of light by dust, salt and other atmospheric aerosols increases the visibility of light
as skyglow while the presence of clouds reflecting light back to earth can substantially
illuminate the landscape (Kyba et al., 2011). Hence the degree of overhead skyglow is a
function of aerosol concentration and cloud height and thickness.
Direct light vs skyglow
Light may appear as either a direct light source from an unshielded lamp with direct line of
sight to the observer, or as skyglow (Figure 20). Skyglow is the diffuse glow caused by source
light that is screened from view, but through reflection and refraction the light creates a glow
in the atmosphere. Skyglow is affected by cloud cover and other particles in the air. Blue light
scatters more in the atmosphere compared to yellow-orange light. Clouds reflect light well,
adding to skyglow.
Figure 20 Skyglow created by lights shielded by a vegetation screen (circled left) and point
sources of light directly visible (circled right).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
32
Distance from light source
The physical properties of light follow the inverse square law which means that the visibility of
the light, as a function of its intensity and spatial extent, decreases with distance from the
source (Figure 21). This is an important factor to consider when modelling light or assessing
the impact of light across different spatial scales, for example across landscape scales
compared to within development footprint.
Figure 21 Modelled changes in the visibility of an unshielded 1,000 W white LED viewed from
A. 10 m; B. 100 m; C. 1 km and D. 3 km.
Measurement of Light
Light has traditionally been measured photometrically or using measurements that are
weighted to the sensitivity of the human eye (peak 555 nm). Photometric light is represented
by the area under the CIE) curve, but this does not capture all radiation visible to wildlife
(Figure 22) (CIE/ISO 23539 Photometry The CIE System of Physical Photometry).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
33
Figure 22 Photometric light represented by the area under the CIE curve (white area) compared
with ability to perceive different wavelengths (black lines) and reported peak sensitivity (black
dots) in humans and wildlife. Note the area under the CIE curve does not include much of the
violet and ultraviolet light visible to many animals. Figure adapted from Campos (2017).
Light can also be measured radiometrically. Radiometric measurements detect and quantify
all wavelengths from the ultraviolet (UV) to infrared (IR). The total energy at every wavelength
is measured. This is a biologically relevant measure for understanding wildlife perception of
light. Terminology, such as radiant flux, radiant intensity, irradiance or radiance all refer to the
measurement of light across all wavelengths of the electromagnetic spectrum.
Understanding the difference between photometry (weighted to the sensitivity of the human
eye) and radiometry (measures all wavelengths) is important when measuring light since many
animals are highly sensitive to light in the blue and the red regions of the spectrum and, unlike
photometry, the study of radiometry includes these wavelengths.
Photometric measures (such as, illuminance and luminance) can be used to discuss the
potential impact of artificial light on wildlife, but their limitations should be acknowledged and
taken into account as these measures may not correctly weight the blue and red wavelengths
to which animals can be sensitive.
Spectral curve
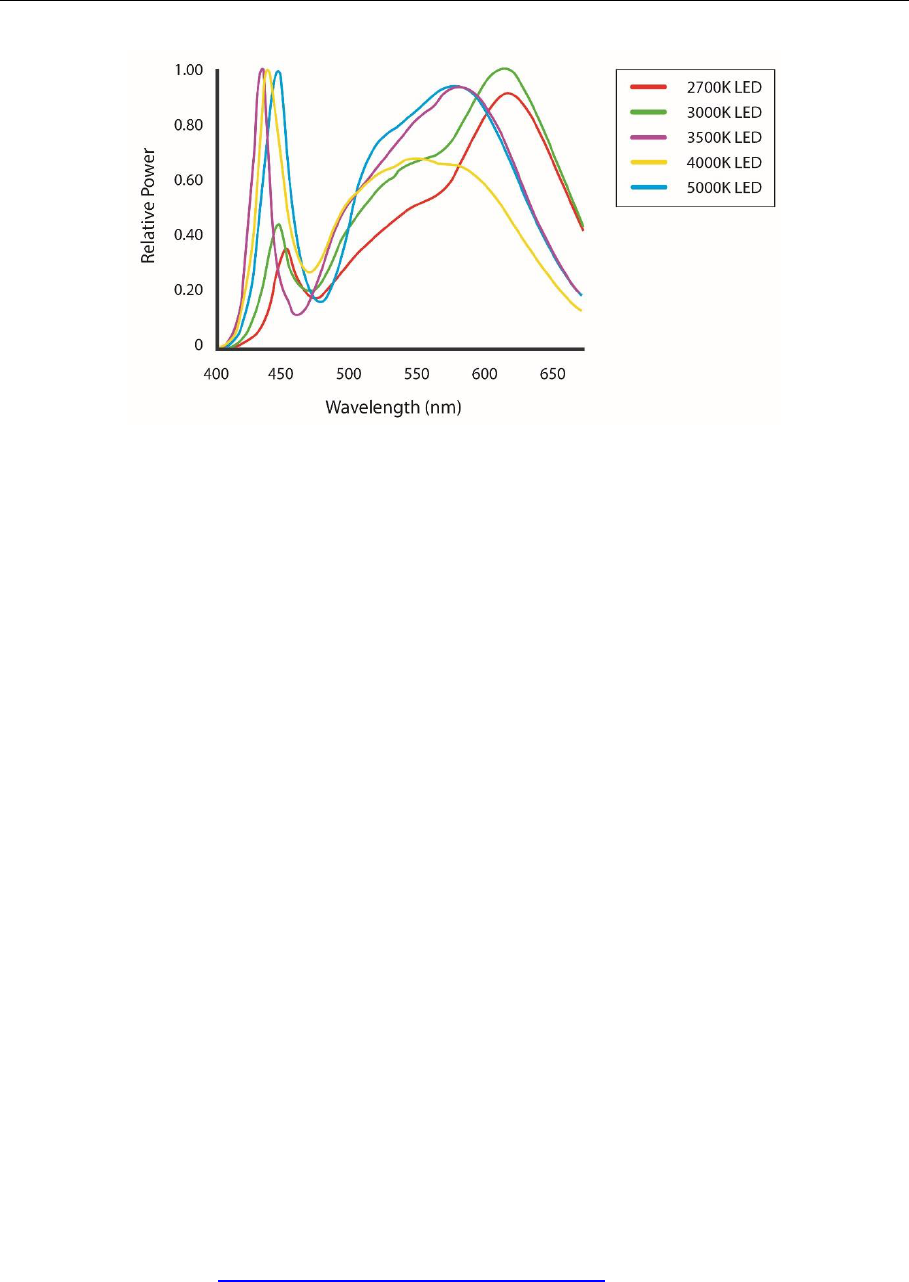
White light is made up of wavelengths of light from across the visible spectrum. A spectral
power curve (Figure 23) provides a representation of the relative presence of each wavelength
emitted from a light source. A lighting design should include spectral power distribution curves
for all planned lighting types as this will provide information about the relative amount of light
emitted at the wavelengths to which wildlife are most susceptible.
UNEP/CMS/COP14/Doc.30.4.4/Annex 3
34
Figure 23 Spectral curves showing the blue content of white 2,700-5,000 K LED lights. Note the
difference in relative power output in the blue (400 - 500 nm) wavelength range. Figure
courtesy of Ian Ashdown.
Light Emitting Diodes (LEDs)
Light emitting diodes are rapidly becoming the most common light type globally as they are
more energy efficient than previous lighting technology. They can be smart controlled, are
highly adaptable in terms of wavelength and intensity, and can be instantly turned on and off.
Characteristics of LED lights that are not found in older types of lamps, but which should be
considered when assessing the impacts of LEDs on wildlife, include:
• With few exceptions, all LED lights contain blue wavelengths (Figure 23 and Figure
24).
• The wattage of an LED is a measure of the electrical energy needed to produce light
and is not a measure of the amount or intensity of light that will be produced by the
lamp.
• The output of light produced by all lamps, including LEDs, is measured in lumens (lm).
• LED lamps require less energy to produce the equivalent amount of light output. For
example, 600 lm output of light requires 40 watts of energy for an incandescent light
bulb and in the year 2020 only 10 watts of energy for a LED lamp. In 2023 less than 5
watts is the input power to achieve 600 lm output. Another way to look at this is that a
100 W incandescent bulb will produce the same amount of light as produced with less
than 10 W from LEDs. Consequently, it is important to not replace an old-style lamp
with the equivalent wattage LED, but to compare the lumen output of the luminaire.
• Different LED lights with the same correlated colour temperature (CCT) can have very
different blue content (Figure 24) yet can appear, to the human eye, to be a similar
colour. As the colour temperature of a white LED increases so can the blue content
(Figure 23). Little or none of this increase in blue wavelength light is measured by
photometric equipment (i.e. lux meter, luminance, illuminance meter, Sky Quality
Meter see Measuring Biologically Relevant Light). LED technology allows for
tuneable RGB colour management. This has the potential to allow for species specific
management of problematic wavelengths (e.g. blue for most wildlife, but also
yellow/orange).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
35
Figure 24 A comparison of the blue wavelength spectral content of two LED lights with the
same CCT (3,500 K). The blue band shows the blue region of the visible spectrum (400–500
nm). The light in A has a much greater blue light content than B yet the two appear to the
human eye as the same colour. For animals with differing sensitivities to light wavelength from
humans, they may appear very different. Figure courtesy of Ian Ashdown.
Correlated colour temperature (CCT)
This describes the colour appearance of a white LED to humans It is expressed in degrees
Kelvin, using the symbol K, which is a unit of measure for absolute temperature. Practically,
a warm yellowish colour have low colour temperatures between 1,000 K and 3,000 K while
lamps characterised by a cool bluish colour have a colour temperature, or CCT, over 5,000 K
(Figure 25). Wavelengths can vary significantly within the same CCT. While lower CCTs are
often recommended, they will not necessarily meet human requirements or mitigate all
impacts. It is important to consider the wildlife impacted and the purpose of the lighting.
Correlated colour temperature does not provide information about the blue content of a lamp.
All LEDs contain blue light (Figure 23) and the blue content generally increases with increased
CCT. The only way to determine whether the spectral content of a light source is appropriate
for use near sensitive wildlife is to consider the spectral curve. For wildlife that is sensitive to
blue light, an LED with low amounts of short wavelength light should be chosen, whereas for
animals sensitive to yellow light (Reed, 1986)
LEDs with little or no light at peak sensitivity
should be used (Longcore et al., 2018).
Figure 25 Correlated colour temperature (CCT) range from warm 1,000 K to cool 10,000 K.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
36
Appendix C - Measuring Biologically
Relevant Light
Animals and humans perceive light differently. Commercial light monitoring
instruments currently focus on measuring the region of the spectrum most visible to
humans. It is important to recognise and account for this fact when monitoring light for
wildlife impact assessment purposes.
Commercial light modelling programmes also focus on light most visible to humans
and this should also be recognised and accounted for in the impact assessment of
artificial light on wildlife.
As different species have different spectral sensitivities to light, there is no general rule
for determining a quantity such as a lux level for illuminance as is done for humans.
For wildlife it is recommended that spectral measurements of irradiance over a wider
range of wavelengths should be made. This range should start at about 300 nm, in the
UV and be extended to the infrared at around 1,000 nm.
Information critical to monitoring the effects of artificial light on wildlife include:
• Spatial extent of skyglow
• Bearings and intensity of light sources along the horizon
• Visibility of light (direct and skyglow) from wildlife habitats
• Spectral distribution of light sources.
Describing the Light Environment
When describing the light environment consideration should be given to how wildlife is likely
to perceive artificial light. Light measurements should be obtained from within important habitat
and taken from a biologically relevant perspective (i.e. close to the ground/from the sky/under
water). Consideration should also be given to elevation from the horizon, the spatial extent of
skyglow and the wavelength distribution (spectrum) of light present.
It is important that light measurements are taken at appropriate times. This may include
biologically relevant times (e.g. when wildlife is using the area). Baseline measurements
should be taken when the moon is not in the sky and when the sky is clear of clouds and in
the absence of temporary lighting (e.g. road works). Conditions should be replicated as closely
as possible for before and after measurements.
Measuring Light for Wildlife
Measuring light to assess its effect on wildlife is challenging and an emerging area of research
and development. Most instruments used to measure skyglow are still in the research phase
with only a few commercial instruments available. Further, the wide range of measurement
systems and units in use globally makes it difficult to choose an appropriate measurement
metric and often results cannot be compared between techniques due to variations in how the
light is measured. There is currently no globally recognised standard method for monitoring
light for wildlife.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
37
Radiometric vs photometric measurement techniques
Radiometric instruments detect and quantify light equally across the spectrum (see
Measurement of Light) and are the most appropriate instruments for monitoring and measuring
light for wildlife management. However, while the techniques to measure radiometric light are
well developed in physics, astronomy and medicine, they are less well developed in
measurement of light in the environment. The instruments currently being developed are
largely the result of academic and/or commercial research and development, are expensive,
and require specialised technical skills for operation, data analysis, interpretation and
equipment maintenance.
The majority of both commercial and research instruments quantify photometric light, which is
weighted to the sensitivity of the human eye, as per the CIE luminosity function curve
described in Measurement of Light. Due to many photometers being modified with filters to
mimic human vision, they do not accurately represent what an animal with high sensitivity to
the blue (400 - 500 nm) or the red (650 - 700 nm) regions of the spectrum will see (Figure 22).
In these cases, the sensitivity to this additional light must be accounted for when reporting
results.
When using photometric instruments for monitoring light this insensitivity to the short and long
wavelength regions of the spectrum should be recognised and accounted for in the
assessment of impact. Information on the spectral power distribution of commercial lights is
readily available from manufacturers and suppliers and should be used to inform any artificial
light impact assessment or monitoring programme. An example of the spectral power
distribution curves for various light sources is shown in Figure 26, along with an overlay of the
CIE curve that represents the light that is measured by all commercial photometric
instruments.
Figure 26 Photometric instruments only quantify light that is within the CIE curve (area under
grey dashed line). This is shown in comparison with the spectral curves of a range of different
light sources.
Recognising that light monitoring instruments for wildlife are in the developmental stage and
that there is a lack of agreed methods and measurement units, monitoring programmes should
aim to measure relevant short and long wavelengths (if possible). The measurement methods
should be clearly described including the region of the spectrum measured, and where not

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
38
measured, how the short and long wavelength regions are being accounted for. Methods to
do this might include a visual assessment of the colour of light in the sky from direct
observation or imagery, where orange glow is typically associated with long wavelength rich
lights (High Pressure Sodium, HPS, Low Pressure Sodium, LPS, PC Amber LED or Amber
LED) and white glow is associated with white light sources rich in short wavelength blue light
(white LEDs, halogens, fluorescents, metal halide etc.).
Alternatively photometric instruments can be used under conditions where the majority of light
sources are the same, for example street lighting or industrial facilities. Monitoring results can
be compared for measurements taken of the same light types (e.g. comparing two HPS
sources, spatially or temporally), but in the context of wildlife monitoring cannot be used to
compare light from an HPS and an LED since they have different wavelength distributions.
This limitation must be taken into account when using photometric instruments to measure
cumulative skyglow, which may include light from multiple sources and light types. Detailed
qualitative spectral information on light types can also be collected to ground truth and confirm
light types contributing to skyglow.
A light monitoring programme might therefore include the collection of a range of different
characteristics of light (e.g. colour, light type, areal extent, spectral power distribution, and
intensity) using various instruments and techniques. These methods and techniques, including
all of the limitations and assumptions, should be clearly stated and considered when
interpreting results. A review of various instrumental techniques for monitoring light is provided
below.
In selecting the most appropriate measuring equipment to monitor the biological impacts of
light on wildlife, it is important to decide what part of the sky is being measured: horizon, zenith
(overhead) or whole sky. For example, marine turtles view light on the horizon between 0° and
30° vertically and integrate across 180° horizontally (Lohmann et al., 1997), so it is important
to include measurement of light in this part of the sky when monitoring for the effects on
hatchling orientation during sea-finding. In contrast, juvenile shearwaters on their first flight
view light in three dimensions (vertically, from below and above) as they ascend into the sky.
Overhead skyglow (zenith) measurements are important when the observer is trying to avoid
glare contamination by point sources of light low on the horizon. Quantifying the whole of
skyglow is important when measuring the effects of cloud cover, which can reflect light back
to illuminate an entire beach, wetland or other habitat.
the cues it uses during orientation, dispersal, foraging, migrating etc. Most wildlife appears to
respond to high intensity short wavelength light, point sources of light, skyglow and directional
light. Consequently, the information likely to be needed to monitor light for wildlife includes:
• The brightness of the entire sky from horizon to horizon.
• The bearing to, intensity of and spectrum of light (point sources and skyglow) on the
horizon. This will dictate the direction in which wildlife can be disoriented.
• The spatial extent of glow near the horizon. A large area of glow on the horizon is likely
to be more visible and disruptive to wildlife than a small area of glow.
• Presence or absence of clouds. Clouds reflect light from distant sources very well,
making an inland source highly visible on the coast, for example. Skyglow is a function
of cloud height, reflectivity and thickness.
• Qualitative information on the light visible to wildlife. An image of light pollution visible
from wildlife habitat can show the spatial extent of light in the sky and direction (see
Figure 20) and in some cases provide information on the light source type (e.g. orange
skyglow will be caused by HPS lights or amber LEDs).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
39
• Emission spectra (colour) of the light. It is particularly important to identify light in the
UV-blue region of the visible spectrum (<500 nm) since this is the light commonly
visible and disruptive to wildlife.
• Also relevant is the maximum brightness under which a light source appears at any
place in the field of view. For example, the full moon creates only an illuminance of
around 0.1 lx, while its luminance is at ~ 2,000 cd/m
2
. This allows some animals to
orient their movement on the direction where the moon appears. Artificial light sources
can exhibit significantly brighter luminance values. In contrast to illuminance,
luminance does not decrease with distance and therefore even bright light sources at
a distance can have attracting effects on wildlife. While luminance is related to human
, is the most
relevant quantity.
Measurement Techniques
Currently, there are no generally agreed methods for measuring biologically relevant light for
wildlife or for quantifying skyglow (Barentine, 2019). This is because most conventional
methods of measuring light are photometric, quantifying only the light under the CIE curve that
is most relevant to the human perception of light. Further, they do not consider the entire night
sky.
There is a need to develop reasonably priced, easily accessible and deployable, repeatable
methods for monitoring biologically relevant light that captures the whole visual field to which
wildlife may be exposed (generally horizon to horizon) (Barentine, 2019). These methods
should be capable of quantifying all wavelengths equally (radiometric) including at least 380
780nm, or capable of being calibrated over the range of wavelengths of relevance for the
species of interest. Optimal methods will have a sensitivity to detect and measure change at
the low light levels represented by artificial light skyglow and must have the ability to
differentiate between individual point sources of light (on a local scale) and skyglow on a
landscape scale (i.e. over tens of kilometres).
It should be noted that measurements needed to assess the impact of skyglow to wildlife may
need to be different from the measurements required to assess light for human safety.
It is anticipated novel methods will be developed with time that will meet the objectives of
monitoring biologically meaningful light and where that occurs, the methods and techniques,
including all of the limitations and assumptions, should be clearly stated for all monitoring
programmes.
Recent reviews have considered various commercial and experimental instrumental
techniques used around the world for quantifying skyglow (Hänel et al., 2018; Barentine 2019).
The reviews assessed the benefits and limitations of the various techniques and made
recommendations for measuring light pollution. Some of these instruments, their benefits and
limitations are discussed below and summarised in Table 1.
Light can be measured in different ways, depending on the objective, landscape scale and
point of view and include:
• remote sensing
• one dimensional (single channel) instruments
• multi-channel instruments
• spectroscopy/spectroradiometry

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
40
Remote sensing
The upward radiance of artificial light at night can be mapped via remote sensing using satellite
or aerial imagery and optical sensors. This information has been used as a socioeconomic
indicator to observe human activity, and increasingly as a tool to consider the impacts of
artificial light on ecosystems (Levin et al., 2020). Examples are:
• The New World Atlas of Artificial Night Sky Brightness
• Light Pollution Map
Benefits: The images are useful as broad scale indicators of light pollution and for targeting
biological and light monitoring programmes. This technique may be a good starting point to
identify potentially problematic areas for wildlife on a regional scale. Images collected via
drones or aircraft may be useful for consideration of artificial light impacts on bird and bat
migrations.
Limitations: Maps derived from satellite collected information have limited value in quantifying
light for wildlife. The images are a measure of light after it has passed though the atmosphere
and been subject to scattering and absorption. They do not give an accurate representation
of the light visible to wildlife at ground level. The annual composite images are made from
images collected under different atmospheric conditions and therefore they cannot be used to
confidently quantify light within or between years. The most commonly used instrument (VIIRS
DNB) is not sensitive to blue light, so light in this part of the spectrum is under sampled. As
satellites with more sophisticated sensors are launched it is expected the value of this
technique to biological monitoring will improve.
Application to wildlife monitoring programmes: Whilst remote sensing tools may provide a
good starting point for identifying artificial light that is problematic for wildlife on a regional
scale, they are currently not an appropriate approach for measuring light as part of a wildlife
monitoring programme as they do not accurately quantify light as observed from the ground,
they underestimate the blue content of light, and results are not repeatable due to
environmental conditions. Images collected via aircraft or drone may have application for
monitoring impacts on airborne wildlife.
One dimensional (single channel) instruments
These instruments measure skyglow using a single channel detector, producing a numerical
value to represent skyglow, typically at the zenith. They are generally portable and easy to
use. They measure skyglow but cannot derive point source information unless they are close
enough such that most of the light detected is emitted from those sources. Examples of single
channel instruments are discussed below.
Sky Quality Meter (SQM)
This is a small handheld unit that quantifies the light in an area of sky (normally directly
overhead at the zenith). Early models had a field of view of around 135° with the more recent
SQM-L model having a narrower 40° diameter field of view. It measures photometric light in
units of magnitudes/arcsec
2
at relatively low detection limits (i.e. it can measure skyglow).
Instrument accuracy is reported at ±10 per cent, although a calibration study on a group of
SQM instruments in 2011 found errors ranging from -16 per cent to +20 per cent (den Outer
et al., 2011). Long term stability of SQMs has not been established.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
41
Reviewers suggest that the first 3-4 measurements from a handheld SQM should be
discarded, then the average of four observations should be collected by rotating the SQM 20°
after each observation to obtain a value from four different compass directions so that the
effects of stray light can be minimised or identified (Hänel et al., 2018). If the measurements
vary by more than 0.2 mag/arcsec
2
the data should be discarded and a new location for
measurements selected. Data should not be collected on moonlit nights to avoid stray light
contaminating the results.
Benefits: The SQM is cheap, easy to use and portable. Some versions have data-logging
capabilities that enable autonomous operation in the field. The sensitivity of the SQM is
sufficient to detect changes in overhead night-time artificial lighting under a clear sky.
Limitations: SQMs cannot be used to resolve individual light sources at a distance, identify
light direction nor can they measure light visible to many wildlife species. The precision and
accuracy of the instrument can vary substantially and an intercalibration study is
recommended to quantify the error of each instrument. Although the SQM is designed to have
a photopic response, it is generally more sensitive to shorter wavelengths (i.e. blue) than a
truly photopic response, but this will depend on the individual instrument. It is not very sensitive
to longer (orange/red) wavelengths (Hänel et al., 2018). The SQM should not be used to
measure light within 20° of the horizon as the detector is designed to measure a homogeneous
sky (such as occurs at the zenith) and does not produce valid data when pointed at a
heterogeneous field of view as observed at the horizon.
Application to wildlife monitoring programmes: A sky quality meter can be used to measure
skyglow directly overhead (zenith) at the wildlife habitat, however, it is important to recognise
its limitations (such as the absence of whole of sky information and inability to measure point
sources of light on the horizon) and follow methods recommended by Hnel et al (2018)
to
ensure repeatability.
Dark Sky Meter
This is an iPhone app that uses the phone camera to collect light and generate a sky
brightness value.
Benefits: It is cheap and easy to use.
Limitations: The Dark Sky Meter is a photometric instrument. It is restricted to Apple iPhones.
It will not work on models older than the 4S and cannot be used to resolve individual lights or
identify light direction. It is relatively imprecise and inaccurate
and cannot reliably measure
light on the horizon (Hänel et al., 2018).
Application to wildlife monitoring programmes: The Dark Sky Meter app is not an appropriate
tool for monitoring light impacts on wildlife as it does not measure biologically relevant light. It
does not provide whole of sky information, it is not able to resolve individual light sources and
it is relatively imprecise and inaccurate. The Dark Sky Meter should be considered more of an
educational tool than a scientific instrument.
Lux Meters and Luminance Meters
Lux meters are commercially available instruments commonly used to measure individual light
sources at close range (i.e. over metres rather than landscape scale). However, the inverse
square law can be used to calculate the illuminance if the distance is known. Lux and
luminance meters measure light based on the photopic sensitivity curve of humans. Lux

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
42
meters measure the light falling on a surface and luminance meters measure the light incident
from a specific solid angle, equivalent to the perceived brightness of the light source.
Benefits: Both can be cheap (with more expensive models available) and easy to use.
Limitations: Both types of devices are photometric, but measurements are weighted to human
perception rather than wildlife. Depending on the sensitivity of equipment, detection limits may
not be low enough to measure typical night sky brightness or illuminance and therefore cannot
measure skyglow for wildlife monitoring purposes. Lux meters have no angular resolution and
luminance meters are coarse so they cannot be used to measure distant light sources at the
horizon precisely.
Application to wildlife monitoring programmes: Commercial lux and luminance meters are not
appropriate for the measurement of light in wildlife monitoring programmes because they have
low sensitivity and low accuracy at low light levels. Expensive tailored devices with enhanced
sensitivity may exist but are still not applicable to wildlife monitoring as they do not measure
biologically relevant light and are not appropriate for use on a landscape scale.
Multi-channel instruments
These instruments map and measure sky brightness by analysing photographic images of the
whole sky. The images are processed to derive a luminance value for all or parts of the sky.
One of the advantages of two-dimensional (wide angle) imaging is that models of natural
sources of light in the night sky can be subtracted from all sky imagery to detect anthropogenic
sources (Duriscoe, 2013). Some examples of devices and techniques to map and measure
night sky brightness using wide-angle images are discussed below.
All-Sky Transmission Monitor (ASTMON)
This charge coupled device (CCD) astronomical camera with fish-eye lens has been modified
by the addition of a filter wheel to allow collection of data through four photometric bands in
the visible spectrum. The spectral range of the instrument is dependent on the sensitivity of
the detector and the filters used but has the advantage of being accurately calibrated on stars.
Benefits: The ASTMON was designed for outdoor installation and the Lite version is portable
with a weather-proof enclosure allowing it to remain outdoors operating robotically for weeks.
It reports data in magnitudes/arcsec
2
for each band and has good precision and accuracy
(Hänel et al., 2018). Once the system is calibrated with standard stars, it can provide
radiometric data for the whole night sky as well as resolve individual light sources.
Limitations: The ASTMON is expensive and requires specialised knowledge to operate and
interpret data. The software provided is not open source and so cannot be modified to suit
individual requirements. The ASTMON may no longer be commercially available. The CCD
cameras used also have a limited dynamic range.
Application to wildlife monitoring programmes: The ASTMON is appropriate for monitoring
artificial light for wildlife as it provides whole night sky measurements that can be calibrated to
give biologically relevant information that is accurate and repeatable.
Digital Camera Equipped with Wide Angle and Fisheye Lenses
This approach is similar to the ASTMON, except using a commercial digital camera with an
RGB matrix rather than a CCD camera with filter wheel, making the system cheaper and more

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
43
transportable. This system provides quantitative data on the luminance of the sky in a single
image (Kolláth, 2010; Jechow et al., 2019).
Benefits: The cameras are easily accessible and portable. When precision is not critical, the
directional distribution of night sky brightness can be obtained. At the very least, the use of a
digital camera with a fisheye lens allows for qualitative imagery data to be collected and stored
for future reference and data analysis. If standard camera settings are used consistently in all
surveys, it is possible to compare images to monitor spatial and temporal changes in sky
brightness. This system also provides multi-colour options with red, green and blue spectral
bands (RGB).
Limitations: Cameras must be calibrated before use and this, together with the specific camera
model, will dictate the precision of the measurements. Calibration for data processing requires
lens vignetting (also known as flat fielding), geometric distortion, colour sensitivity of the
camera, and sensitivity function of the camera. Specialised knowledge is required to process
and interpret these images. Also, like CCD cameras, the detectors in digital cameras have a
limited dynamic range which can easily saturate in bright environments. In addition, fisheye
systems often produce the poorest quality data at the horizon where the distortion due to the
lens is the greatest.
Calibrating the camera is difficult and standard methods have not been developed. Laboratory
or astronomical photometric techniques are generally used which require specialist knowledge
and expertise. A precision of ~10 per cent can be achieved using this technique. Standard
commercial cameras are calibrated to the human eye (e.g. photometric), however, the ability
to obtain and process an image allows for qualitative assessment of light types (based on the
colour of skyglow), which provides additional data for interpreting the biological relevance of
the light.
Application to wildlife monitoring programmes: A digital camera equipped with wide angle or
fisheye lenses is appropriate for measuring light in wildlife monitoring programmes as it
provides horizon to horizon information with enough sensitivity and accuracy to detect
significant changes in low light environments. Images allow for detection of both skyglow, light
source type, and point source information. When data is manually processed biologically
relevant measurements can be obtained. Because the system is fast, dynamics of skyglow
and direct light can be monitored (Jechow et al., 2018).
All Sky Mosaics
This technique was developed by the US National Parks Service and provides an image of
the whole of the sky by mosaicking 45 individual images. The system comprises a CCD
camera, a standard 50 mm lens, an astronomical photometric Bessel V filter with IR blocker
and a computer controlled robotic telescope mount. Data collection is managed using a
portable computer, commercial software and custom scripts.
Benefits: The angular resolution, precision and accuracy of the system is good, and it is
calibrated and standardised on stars. The images produced have high resolution. The system
is best suited for long term monitoring from dark sky sites. However, with the addition of a
neutral density filter, the luminance or illuminance of a near-by bright light source can be
measured. Also, other photometric bands can be measured with the use of additional filters.
Limitations: The system is expensive and requires specialised knowledge to operate the
system, analyse and interpret the data. These cameras are calibrated to the human eye with
the inclusion of a visible filter, however the ability to obtain and process an image allows for
qualitative assessment of light types in the (based on the colour of skyglow), which provides

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
44
additional data for interpreting the biological relevance of the light. Measurement procedures
are time consuming and require perfect clear sky conditions and single spectral band, or
repeated measurements are required.
Application to wildlife monitoring programmes: All sky mosaics would be an appropriate tool
for monitoring of artificial light for wildlife. They provide whole of sky images with high
resolution and, with appropriate filters, can be used to measure biologically relevant
wavelength regions.
Mobile luminance cameras
Benefits: New and affordable mobile luminance cameras are able to produce high resolution
false-colour images of the measured surrounding in high optical resolution like a photograph
and include software for evaluation. The camera is based on a DSLR and can be used to
measure very low light levels. Due to the photographic image resolution and assessment,
multiple light sources do not overlap and can be assessed simultaneously even if they are
next to each other. Luminance values are calculated from numerical transformations of RGB
sensor data. This can be an effective way of characterising light fields in the night-time
environment if 1) the data are use appropriately and in the correct units; and 2) instruments
are properly calibrated for use in typical outdoor night-time lighting levels.
Limitations: Mobile luminance cameras are still related to human sensitivity. The luminance is
calculated from an RGB-image in RAW format (i.e. digital image file). UV and IR cannot be
assessed by these devices. Images are taken with standard camera sensitivities and require
higher light levels at the target area of the photograph and so are not suitable to assess low
level disturbances like skyglow.
Application to wildlife monitoring programmes: Mobile luminance cameras could be used to
assess potentially disturbing light sources.
Spectroscopy/spectroradiometry
Different light types produce a specific spectral signature or spectral power distribution (for
example Figure 26). Using a spectrometer it is possible to separate total sky radiance into its
contributing sources based on their spectral characteristics. Being able to assess the impacts
of different light sources is of relevance during this time of transition in lighting technology.
Where wildlife sensitivity to particular wavelength regions of light is known, being able to
capture the spectral power distributions of artificial light and then predict how the light will be
perceived by wildlife will be of particular benefit in assessing the likely impacts of artificial light.
This type of approach has been utilised in astronomy for a long time, but only recently applied
to measurement and characterisation of light pollution on earth. An example of a field
deployable spectrometer - the Spectrometer for Aerosol Night Detection (SAND) is described
below.
Spectrometer for Aerosol Night Detection (SAND)
SAND uses a CCD imaging camera as a light sensor coupled with a long slit spectrometer.
The system has a spectral range from 400 nm to 720 nm and is fully automated. It can
separate sampled sky radiance into its major contributing sources.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
45
Benefits: This approach can quantify light at specific wavelengths across the spectrum
different light types.
Limitations: Calibration, collection and interpretation of these data requires specialist
knowledge and equipment and is expensive. SAND does not provide whole sky information.
Application to wildlife monitoring programmes: The use of a portable spectrometer that can
identify light types based on their spectral power distribution or measure light at specific
wavelengths of interest would be a useful contribution to a wildlife monitoring programme.
Unfortunately, the prototype SAND instrument is no longer in operation. However, this
instrument exemplifies the type of approaches that will be of benefit for measuring light for
wildlife in the future.
Most appropriate instrument for measuring biologically relevant light
The most appropriate method for measuring light for wildlife will depend on the species present
and the type of information required. In general, an appropriate approach will quantify light
across the whole sky, across all spectral regions, differentiating point light sources from
skyglow and it will be repeatable and easy to use.
The digital camera and fisheye lens technique was recommended by Hnel et al. (2018) and
Barentine (2019) as the best compromise between cost, ease-of-use and amount of
information obtained when measuring and monitoring skyglow. Hnel et al. (2018) did,
however, recognise the urgent need for the development of standard software for calibration
and displaying results from light monitoring instruments. In the future, hyperspectral cameras
with wide field of view might become available combining the advantages of spectroradiometry
and all-sky imagery. However, such devices do not currently exist.
It should be noted that this field is in a stage of rapid development.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
46
Table 1 Examples of instrumental light measurement techniques (modified from Hnel et al., 2018).
Instrument
Measurement
Units
Detect
SkyGlow
Data
Type
Spectrum
measured
Scale
Commercially
Available
Data
Quality
Cost
Remote sensing:
Satellite imagery
Various
Yes via
modelling
Images +
numerical
value
Single band
Landscape
Yes
Mod-
high
Some datasets
free
One dimensional:
Sky Quality Meter
(SQM)
magSQM/arcsec
2
Yes
Numerical
value
Single band
Overhead
Yes
Mod
Low cost
Dark Sky Meter
(iPhone)
~
magSQM/arcsec
2
Yes
Numerical
value
Single band
Overhead
Yes
Low
No cost /
negligible
Luxmeter
lux
No
Numerical
value
Single band
Metres
Yes
Low
Low cost
Multi channel:
ASTMON
mag
v
/arcsec
2
Yes
Image +
numerical
value
Multi band
filter wheel
Whole sky
No
High
High cost
DSLR + fisheye
~cd/m
2
,
~mag
v
/arcsec
2
Yes
Image +
numerical
value
Multi band
RGB
Whole sky
Yes
Mod-
high
Medium cost
All sky mosaic
cd/m
2
,
mag
v
/arcsec
2
Yes
Image +
numerical
value
Single band
Whole sky
No
High
High cost
Mobile luminance
camera
cd/m
2
No
Image +
numerical
value
RGB sensor
Landscape
Disturbing lights
Yes
High
High cost
Spectroradiometry:
Spectrometer for
Aerosol Night
Detection (SAND)
W/(m
2
nm sr)
Yes
Spectral
power
curve
Multi band
hyperspectral
Landscape
No
Mod-
high
High cost

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
47
Modelling Predicted Light
Available commercial light models
Most modelling software that is currently available is problematic as the models are weighted
towards a human perception of light as represented by the CIE/photometric curve and do not
account for the wavelengths to which wildlife are most sensitive. For example, most wildlife is
sensitive to short wavelength violet and blue light (Figure 17), but little or none of this light is
measured by commercial instruments and consequently it is not accounted for in current light
models.
A second limitation of many light models for biology is the inability to accurately account for
environmental factors, such as: atmospheric conditions (moisture, cloud, rain, dust); site
topography (hills, sand dunes, beach orientation, vegetation, buildings); other natural sources
of light (moon and stars); other artificial sources of light; the spectral output of luminaires; and
the distance, elevation, and viewing angle of the observing species. Such a model would
involve a level of complexity that science and technology has yet to deliver.
A final major limitation is the lack of biological data with which to confidently interpret a model
outcome. Therefore, it is not possible to objectively estimate how much artificial light is going
to cause an impact on a particular species, or age class, over a given distance and under
variable environmental conditions.
Recognising these limitations, it can still be valuable to model light during the design phase of
new lighting installations to test assumptions about the light environment. For example,
models could test for the potential for light spill and line of sight visibility of a source. These
assumptions should be confirmed after construction.
Development of modelling tools that can take account of broad spectral data and
environmental conditions are in the early stages of development but are rapidly improving
(Barentine, 2019).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
48
Appendix D Artificial Light Auditing
Industry best practice requires onsite inspection of a build to ensure it meets design
specifications. An artificial light audit should be undertaken after construction to confirm
compliance with the artificial lighting management plan.
An artificial light audit cannot be done by modelling of the as-built design alone and should
include a site visit to:
• Confirm compliance with the artificial lighting management plan
• Check as-built compliance with engineering design
• Gather details on each luminaire in place
• Conduct a visual inspection of the facility lighting from the wildlife habitat
• Review the artificial light monitoring at the project site
• Review artificial light monitoring at the wildlife habitat.
Following completion of a new project or modification/upgrade of the lighting system of an
existing project, the project should be audited to confirm compliance with the artificial lighting
management plan.
Step-by-Step Guide
The steps to carry out an artificial light audit include:
• Review of the artificial lighting management plan
• Review of best practice light management or approval conditions
• Review of as-built drawings for the lighting design
• Check for compliance with the approved pre-construction (front end) lighting design
• Conduct a site inspection both during the day and at night to visually check and
measure the placement, number, intensity, spectral power output, orientation, and
management of each lamp and lamp type. Where possible this should be done with
the lighting in operation and with all lighting extinguished.
• Measurements should be taken in a biologically meaningful way. Where there are
limitations in measurements for wildlife these should be acknowledged.
• Record, collate and report on the findings and include any non-conformances. This
should consider any differences between baseline and post construction observations.
Where lighting outputs were modelled as part of the design phase, actual output should
be compared with modelled scenarios.
• Make recommendations for any improvements or modifications to the lighting design
that will decrease the impact on wildlife.
The audit should be conducted by an appropriately qualified environmental
practitioner/technical specialist during a site visit. The audit should also include:
• A visual inspection of the facility lighting from the location of the wildlife habitat and
where feasible the perspective of the wildlife (i.e. sand level for a marine turtle)
• Artificial light monitoring at the project site
• Artificial light monitoring at the wildlife habitat.
A post-construction site visit is critical to ensure no previously unidentified lighting issues are
overlooked.
UNEP/CMS/COP14/Doc.30.4.4/Annex 3
49
Appendix E Artificial Light Management Checklist
Table 2 provides a checklist of issues to be considered during the environmental assessment of new infrastructure involving artificial light, or
upgrades to existing artificial lighting for both proponents and assessors. Table 3 provides a checklist of issues to be considered for existing
infrastructure with external lighting where species are observed to be impacted by artificial light. Relevant sections of the Guidelines are provided
for each issue.
Table 2 Checklist for new developments or lighting upgrades.
Issue to be considered
Light owner or manager
Regulator
Further information
Pre-development
What are the regulatory
requirements for artificial light
for this project?
Is an EIA required? What other
requirements need to be addressed?
What information should be
sought from the proponent as part
of the assessment process?
Regulatory considerations for
the management of artificial
light
Does the lighting design follow
principles of best practice?
What is the purpose of the artificial
light for this project?
Does the project use the
principles of best practice light
design?
Best practice light design
What wildlife is likely to be
affected by artificial light?
Review species information within 20
km of the proposed development.
Assess species information
Wildlife and artificial light
What light management and
impact mitigation will be
implemented?
What light mitigation and
management will be most effective
for the affected species?
Is the proposed management and
mitigation likely to reduce the
effect on species of conservation
concern?
Species specific technical
appendices and species
expert guidance
How will light be modelled?
Is light modelling appropriate? How
will the model be used to inform light
management for wildlife?
Are the limitations of light
modelling for wildlife appropriately
acknowledged?
Modelling predicted light
Have all lighting-relevant
considerations been included in
the lighting management plan?
Have all steps in the EIA process
been undertaken and documented in
the lighting management plan?
Does the lighting management
plan comprehensively describe all
steps in the EIA process?
Environmental impact
assessment for effects of
artificial light on wildlife
Lighting Management Plan
How will continuous
improvement be achieved?
How will light management be
evaluated and adapted?
Is a continuous review and
improvement process described?
Lighting Management Plan

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
50
Issue to be considered
Light owner or manager
Regulator
Further information
Post development
How will lighting be measured?
What is the most appropriate
technique(s) for measuring
biologically relevant light and what
are the limitations?
Ensure appropriate light
measurement techniques are
used and limitations of the
methods recognised
Measuring biologically
relevant light
How will lighting be audited?
What is the frequency and
framework for in-house light
auditing?
How will the results of light audits
feed back into a continuous
improvement process?
Artificial light auditing
Is artificial light affecting wildlife?
Does the biological monitoring
indicate an effect of artificial light on
wildlife and what changes will be
made to mitigate this impact?
Is there a process for addressing
monitoring results that indicate
there is a detectable light impact
on wildlife, and is it appropriate?
Wildlife and artificial light
Lighting Management Plan
Managing existing light
pollution
What adaptive management can
be introduced?
How will the results of light audits
and biological monitoring be used in
an adaptive management
framework, and how will
technological developments be
incorporated into artificial light
management?
What conditions can be put in
place to ensure a continuous
improvement approach to light
management?
Lighting Management Plan

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
51
Table 3 Checklist for existing infrastructure
Consideration
Light owner or manager
Regulator
Further information
Is wildlife exhibiting a change in
survivorship, behaviour or
reproduction that can be
attributed to artificial light?
What species of conservation
concern are found within 20 km
of light source? Are there dead
animals or are animals displaying
behaviour consistent with the
effects of artificial light?
Is there evidence to implicate
artificial light as the cause of the
change in wildlife survivorship,
behaviour or reproductive
output?
Review existing environmental
approvals
Describe wildlife
Wildlife and artificial light
Regulatory considerations for the
management of light
Species expert advice
Is lighting in the area best
practice?
Are there modifications or
technological upgrades that
could be made to improve
artificial light management?
Are there individual light owners
or managers who can be
approached to modify current
lighting?
Principles of best practice lighting
design
Is the light affecting wildlife from
a single source or multiple
sources?
Are there multiple stakeholders
that need to come together to
address the cumulative light
pollution?
Is there a role for government to
facilitate collaboration between
light owners and managers to
address light pollution?
Managing existing light pollution
Lighting Management Plan
Can appropriate monitoring be
undertaken to confirm the role of
artificial light in wildlife
survivorship, behavioural or
reproductive output changes?
How much light is emitted from
the property and is it affecting
wildlife?
Facilitate wildlife monitoring.
Field surveys for wildlife
Measuring biologically relevant
light
Species expert advice
How will artificial light be
audited?
What is the frequency and
framework for in-house light
auditing?
Can a light audit be undertaken
on a regional scale?
Artificial light auditing
What adaptive light management
can be introduced?
Are there improvements in
lighting technology that can be
incorporated into existing
lighting?
What changes can be
implemented in response to
biological monitoring and light
audits?
Specialist lighting engineer
advice

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
52
Appendix F - Marine Turtles
Marine turtles nest on sandy beaches. There is a robust body of evidence
demonstrating the effects of light on turtle behaviour and survivorship. Light is likely
to affect turtles if it can be seen from the nesting beach, nearshore or adjacent waters.
Adult females may be deterred from nesting where artificial light is visible on a
nesting beach. Hatchlings may become misoriented or disoriented and be unable to
find the sea or successfully disperse to the open ocean. The effect of light on turtle
behaviour has been observed from lights up to 18 km away.
The physical aspects of light that have the greatest effect on turtles include intensity,
colour (wavelength), and elevation above beach. Management of these aspects will
help reduce the threat from artificial light.
Seven species of marine turtles are listed on the CMS appendices: the green (Chelonia
mydas), loggerhead (Caretta caretta), hawksbill (Eretmochelys imbricata), olive ridley
(Lepidochelys olivacea ridley (Lepidochelys kempii), flatback (Natator depressus)
and leatherback (Dermochelys coriacea) turtles. Artificial light can disrupt critical behaviours
such as adult nesting and hatchling orientation, sea-finding and dispersal, and can reduce the
reproductive viability of turtle populations.
Figure 27 Loggerhead turtle (Caretta caretta). Photo: David Harasti.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
53
Conservation Status
Marine turtles are protected under international treaties and agreements including the
Convention on the Conservation of Migratory Species of Wild Animals (CMS, Bonn 1979), the
Convention on International Trade in Endangered Species of Flora and Fauna (CITES,
Washington 1973), the CMS Memorandum of Understanding concerning Conservation
Measures for Marine Turtles of the Atlantic Coast of Africa (1999), the CMS Memorandum of
Understanding on the Conservation and Management of Marine Turtles and their Habitats of
the Indian Ocean and South-East Asia (IOSEA, 2001) and the Inter-American Convention for
the Protection and Conservation of Sea Turtles (2001). CMS adopted the Single Species
Action Plan for Loggerhead Turtle (Caretta caretta) in the South Pacific Ocean in 2014, and
the Single Species Action Plan for the Hawksbill Turtle (Eretmochelys imbricata) in South-East
Asia and the Western Pacific Ocean Region in 2022. See Table 4 for CMS listings and IUCN
Red List statuses of marine turtle species.
Table 4: Marine turtle listings on CMS appendices and IUCN Red List Statuses (CMS, 2023a;
IUCN, 2023)
Common name
Scientific name
Year of
Appendix I
Listing
Year of
Appendix II
Listing
Global IUCN Red
List Status and
Trend (2023)
Green Turtle
Chelonia mydas
1979
1979
Endangered
(decreasing)
Loggerhead
Turtle
Caretta caretta
1985
1979
Vulnerable
(decreasing)
Hawksbill Turtle
Eretmochelys
imbricata
1985
1979
Critically
Endangered
(decreasing)
Turtle
Lepidochelys
kempii
1979
1979
Critically
Endangered
(unknown)
Olive Ridley
Turtle
Lepidochelys
olivacea
1985
1979
Vulnerable
(decreasing)
Leatherback
Turtle
Dermochelys
coriacea
1979
1979
Vulnerable
(decreasing)
Flatback Turtle
Natator depressus
-
1979
Data Deficient
Distribution and Habitat
Turtle nesting habitats include sub-tropical and tropical beaches. Each nesting population is
different according to local conditions and, therefore, sensitive times such as peak nesting
periods need to be determined on a case-by-case basis for management to be effective. The
effect of artificial lights on turtles is most pronounced at nesting beaches and in nearshore
waters, which might include inter-nesting areas, through which hatchlings travel to reach the
ocean.
The IUCN-SSC Marine Turtle Specialist Group has developed a set of criteria and a framework
for defining Important Marine Turtle Areas (IMTAs). IMTAs are
marine turtle regional management units (RMUs) that are of particular biological significance
for the persistence of marine turtles, and/or where the contributions of marine turtles to
traditions and cultures of local peopl

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
54
Effects of Artificial Light on Marine Turtles
The effect of artificial light on turtle behaviour has been recognised since 1911(Hooker, 1911).
Since then a substantial body of research has focused on how light affects turtles and its effect
on turtle populations (Witherington and Martin, 2003; Lohmann et al.,1997; Salmon, 2003).
The global increase in light pollution from urbanisation and coastal development (Falchi et al.,
2016)
is of particular concern for turtles since their important nesting habitat frequently
overlaps with areas of large-scale urban and industrial development, which have the potential
to emit a large amount of light, including direct light, reflected light, skyglow and gas flares
(Pendoley, 2000; Pendoley, 2005; Kamrowski et al., 2012).
Effect of artificial light on nesting turtles
Although they spend most of their lives in the ocean, females nest on sandy tropical and
subtropical beaches, predominantly at night. They rely on visual cues to select nesting
beaches and orient on land. Artificial night lighting on or near beaches has been shown to
disrupt nesting behaviour (Witherington and Martin, 2003). Beaches with artificial light, such
as urban developments, roadways, and piers typically have lower densities of nesting females
than dark beaches (Salmon, 2003; Hu et al., 2018).
Some light types do not appear to affect nesting densities (Low Pressure Sodium, LPS, and
filtered High Pressure Sodium, HPS, which excludes wavelengths below 540 nm)
(Witherington, 1992; Pennell, 2000). On beaches exposed to light, females will nest in higher
numbers in areas that are shadowed (Price et al., 2018; Salmon et al., 1995). Moving sources
of artificial light may also deter nesting or cause disturbance to nesting females (e.g. flash
photography) (Campbell, 1994).
Effect of artificial light on hatchlings emerging from the nest
Most hatchling turtles emerge at night
and must rapidly reach the ocean to avoid predation
(Mrosovsky, 1968; Erb and Wyneken, 2019). Hatchlings locate the ocean using a combination
of topographic and brightness cues, orienting towards the lower, brighter oceanic horizon and
away from elevated darkened silhouettes of dunes and/or vegetation behind the beach
(Lohmann et al., 1997; Limpus and Kamrowski, 2013; Pendoley and Kamrowski, 2015a). They
can also find the sea using secondary cues such as beach slope (Lohmann et al., 1997).
Sea-finding behaviour may be disrupted by artificial lights, including flares, which interfere with
natural lighting and silhouettes (Pendoley 2000; Witherington and Martin, 2003; Kamrowski et
al., 2014; Pendoley and Kamrowski, 2015a). Artificial lighting may adversely affect hatchling
sea-finding behaviour in two ways: disorientation - where hatchlings crawl on circuitous paths;
or misorientation - where they move in the wrong direction, possibly attracted to artificial lights
(Witherington and Martin, 2003; Salmon, 2006). On land, movement of hatchlings in a direction
other than the sea often leads to death from predation, exhaustion, dehydration, or being
crushed by vehicles on roads (Erb and Wyneken, 2019).
Wavelength, intensity and direction
Brightness is recognised as an important cue for hatchlings as they attempt to orient toward
the ocean. Brightness refers to the intensity and wavelength of light relative to the spectral
sensitivity of the receiving eye (Witherington and Martin, 2003). Both field and laboratory-
based studies indicate that hatchlings have a strong tendency to orient towards the brightest
direction. The brightest direction on a naturally dark beach is typically towards the ocean

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
55
where the horizon is open and unhindered by dune or vegetation shadows (Limpus and
Kamrowski, 2013).
The attractiveness of hatchlings to light differs by species but, in general, artificial lights most
disruptive to hatchlings are those rich in short wavelength blue and green light (e.g. metal
halide, mercury vapour, fluorescent and LED) and lights least disruptive are those emitting
long wavelength pure yellow-orange light (e.g. high or low pressure sodium vapour)
(Witherington and Bjorndal, 1991; Pendoley, 2005; Horch et al., 2008; Fritches, 2012).
Loggerhead turtles are particularly attracted to light at 580 nm, green and flatback turtles are
attracted to light <600 nm with a preference to shorter wavelength light over longer wavelength
light, and many species are also attracted to light in the ultraviolet range (<380 nm)
(Witherington and Bjorndal, 1991; Levenson et al., 2004; Pendoley, 2005; Fritches, 2012).
Although longer wavelengths are less attractive than shorter wavelengths, they can still disrupt
sea-finding, and if bright enough can elicit a similar response to shorter wavelength light
(Mrosovsky and Shettleworth, 1968; Mrosovsky, 1972; Pendoley, 2005; Pendoley and
Kamrowski, 2015ab; Robertson et al., 2016). Hence, the disruptive effect of light on hatchlings
is also strongly correlated with intensity. Red light must be almost 600 times more intense than
blue light before green turtle hatchlings show an equal preference for the two colours
(Mrosovsky, 1972). It is therefore important to consider both the wavelength and the intensity
of the light.
Since the sun or moon may rise behind the dunes on some nesting beaches, hatchlings
attracted to these point sources of light would fail to reach the ocean. Hatchlings orientate
themselves by integrating light across a horizontally broad (180° for green, olive ridley and
-
s that
light closest to the horizon plays the greatest role in determining orientation direction, so it is
important to consider the type and direction of light that reaches the hatchling (Lohmann et
al., 1997).
As a result of these sensitivities, hatchlings have been observed to respond to artificial light
up to 18 km away during sea-finding (Kamrowski et al., 2014).
Shape and form
Horizon brightness and elevation are also important cues for hatchling orientation. In
laboratory and field studies hatchlings move away from elevated dark horizons and towards
the lowest bright horizon (Limpus and Kamrowski, 2013; Salmon et al., 1992). However, in
situations where both cues are present, hatchlings are more responsive to the effects of
silhouettes and darkened horizon elevation than to differences in brightness. On a natural
beach this behaviour would direct the hatchlings away from dunes and vegetation and towards
the more open horizon over the ocean.
This hypothesis has been supported by field experiments where hatchling sea-finding was
significantly less ocean oriented when exposed to light at 2° elevation compared with 16°
elevation, emphasising the importance of horizon elevation cues in hatchling sea-finding
(Pendoley and Kamrowski, 2015a).
Effect of artificial light on hatchlings in nearshore waters
Artificial lights can also interfere with the in-water dispersal of hatchlings (Witherington and
Bjorndal, 1991). Hatchlings leaving lit beaches spend longer crossing nearshore waters and
can be attracted back to shore (Harewood and Horrocks, 2008). A study in Costa Rica found

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
56
that olive ridley turtles were still attracted to lights when they were in the ocean (Cruz et al.,
2018). This has implications for any attempts to mitigate the negative impact of artificial light
in habitat used by turtles. At sea, hatchlings have been reported swimming around lights on
boats (Limpus et al., 2003; White and Gill, 2007)
and in laboratory studies lights have attracted
swimming hatchlings (Salmon and Wyneken, 1990). Recent advances in acoustic telemetry
technology have allowed hatchlings to be passively tracked at sea, demonstrating that
hatchlings are attracted to lights at sea and spend longer in the nearshore environment when
lights are present (Thums et al., 2016; Wilson et al., 2018). This attraction can divert hatchlings
from their usual dispersal pathway, causing them to linger around a light source, or become
trapped in the light spill (Wilson et al., 2018). Hatchlings actively swim against currents to
reach light, which is likely to reduce survival either from exhaustion and/or predation. An
additional problem is that light sources are associated with structures that also attract fish
(such as jetties), so there will be increased predation (Wilson et al., 2019).
Environmental Impact Assessment of Artificial Light on
Marine Turtles
Infrastructure with artificial lighting that is externally visible should implement Best Practice
Lighting Design as a minimum. Where there is important habitat for turtles within 20 km of a
project, an EIA should be undertaken. The following sections step through the EIA process
with specific consideration for turtles.
The 20 km buffer for considering important habitat is based on skyglow approximately 15 km
from the nesting beach affecting flatback hatchling behaviour
and light from an aluminium
refinery disrupting turtle orientation 18 km away (Kamrowski et al., 2014; Hodge et al., 2007).
Where artificial light is likely to influence marine turtle behaviour, consideration should be given
inform the design phase.
The presence of boats with artificial lights should be taken into consideration, especially when
neonates are hatching.
Associated guidance
• Single Species Action Plan for the Loggerhead Turtle (Caretta caretta) in the South
Pacific Ocean
• Single Species Action Plan for the Hawksbill Turtle (Eretmochelys imbricata) in
South-East Asia and the Western Pacific Ocean Region
• IOSEA (Memorandum of Understanding on the Conservation and Management of
Marine Turtles and their Habitats of the Indian Ocean and South-East Asia)
Conservation and Management Plan
• IOSEA Guidelines for the review EIAs of developments impacting on sea turtles and
turtle habitat
• Inter-American Convention for the Protection and Conservation of Sea Turtles
website
• The State of the World’s Sea Turtles (SWOT) Report, Vol. XVIII
Qualified personnel
Lighting design/management and the EIA process should be undertaken by appropriately
qualified personnel. Lighting management plans should be developed and reviewed by

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
57
appropriately qualified lighting practitioners who should consult with an appropriately qualified
marine biologist or ecologist.
Step 1: Describe the project lighting
Information collated during this step should consider the Effects of Artificial Light on Marine
Turtles. Turtles are susceptible to the effect of light on beaches and in the water, so the
location and light source (both direct and skyglow) should be considered. Turtles are most
sensitive to short wavelength (blue/green) light and high intensity light of all wavelengths.
Hatchlings are most susceptible to light low on the horizon. They orient away from tall dark
horizons so the presence of dunes and/or a vegetation buffer behind the beach should be
considered at the design phase.
Step 2: Describe marine turtle population and behaviour
The species and the genetic population nesting in the area of interest should be described.
This should include the conservation status of the species; population trends (where known);
how widespread/localised nesting for that population is; the abundance of turtles nesting at
the location; the regional importance of this nesting beach; and the seasonality of
nesting/hatching.
Where there is insufficient data to understand the population importance or demographics, or
where it is necessary to document existing turtle behaviour, field surveys and biological
monitoring may be necessary.
Biological monitoring of marine turtles
Any monitoring associated with a project should be developed, overseen and results
interpreted by appropriately qualified personnel to ensure reliability of the data.
The objectives of turtle monitoring in an area likely to be affected by artificial light include:
• understanding the size and importance of the population;
• describing turtle behaviour before the introduction/upgrade of light; and
• assessing nesting and hatchling orientation behaviour to determine the cause of any
existing or future misorientation or disorientation.
The data will be used to inform the EIA and assess whether mitigation measures are
successful. Suggested minimum monitoring parameters (what is measured) and techniques
(how to measure them) are summarised in Table 5.
As a minimum, qualitative descriptive data on visible light types, location and directivity should
also be collected at the same time as the biological data. Handheld-camera images can help
describe the light. Quantitative data on existing skyglow should be collected, if possible, in a
biologically meaningful way, recognising the technical difficulties in obtaining these data. See
Measuring Biologically Relevant Light for a review.
Table 5 Recommended minimum biological information necessary to assess the importance of
a marine turtle population and existing behaviour, noting that the risk assessment will guide the
extent of monitoring (e.g. a large source of light visible over a broad spatial scale will require
monitoring of multiple sites whereas a smaller localised source of light may require fewer sites
to be monitored).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
58
Target Age
Class
Survey Effort
Duration
Reference
Adult Nesting
Daily track census over 11.5
internesting cycles at peak
of
the nesting season (1421
days).
If the peak nesting period for
this population/at this location
has not been defined, then a
study should be designed in
consultation with a qualified
turtle biologist to determine
the temporal extent of activity
(i.e. systematic monthly
surveys over a 12-month
period).
Minimum
two breeding
seasons
Eckert et al. (1999)
Pendoley et al.
(2016)
Queensland
Marine Turtle Field
Guide
North West Shelf
Flatback Turtle
Conservation Plan
Turtle Monitoring
Field Guide
SWOT Minimum
Data Standards for
Sea Turtle Nesting
Beach Monitoring
Research and
Management
Techniques for the
Conservation of
Sea Turtles
Hatchling
Orientation
Minimum of 14 days over a
new moon phase about 50
days* after the peak of adult
nesting.
Beach: Hatchling fan
monitoring.
In water: Hatchling tracking
Minimum two
breeding seasons
Pendoley (2005)
Kamrowski et al.
(2014)
Witherington
(1997)
Thums et al.
(2016)
*Incubation time will be population specific.
To understand existing hatchling behaviour, it will be necessary to undertake monitoring (or
similar approach) to determine hatchling ability to locate the ocean and orient offshore prior to
construction/lighting upgrades.
A well-designed monitoring programme will capture:
• hatchling behaviour
at the light exposed beach and a control/reference beach
(Witherington, 1997; Pendoley, 2005; Kamrowski et al., 2014);
• hatchling behaviour before project construction begins to establish a benchmark to
measure against possible changes during construction and operations;

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
59
• hatchling behaviour on a new moon to reduce the influence of moonlight and capture
any worst case scenario effects of artificial light on hatchling orientation;
• hatchling behaviour on full moon nights to assess the relative contribution of the
artificial light to the existing illuminated night sky.
Ideally, survey design will have been set up by a quantitative ecologist/biostatistician to ensure
that the data collected provides for meaningful analysis and interpretation of findings.
Step 3: Risk assessment
Management of artificial light should ensure turtles are not displaced from important habitat
and that anthropogenic activities in important habitat are managed so that biologically
important behaviour can continue. These consequences should be considered in the risk
assessment process. The aim of these Guidelines is that light is managed to ensure that at
important nesting beaches females continue to nest on the beach, post-nesting females return
to the ocean successfully, emerging hatchlings orient in a seaward direction and dispersing
hatchlings can orient successfully offshore.
Consideration should be given to the relative importance of the site for nesting. For example,
if this is the only site at which a population nests, a higher consequence rating should result
from the effects of artificial light.
In considering the likely effect of light on turtles, the risk assessment should consider the
existing light environment, the proposed lighting design and mitigation/management, and the
behaviour of turtles at the location. Consideration should be given to how the turtles will
perceive light. This should include wavelength and intensity information as well as perspective.
To assess how/whether turtles are likely to see light, a site visit should be made at night and
the area viewed from the beach (approximately 10 cm above the sand) as this will be the
perspective of the nesting turtles and emerging hatchlings. Similarly, consideration should be
given to how turtles (both adults and hatchlings) will see light when in nearshore water.
Using this perspective, the type and number of lights should be considered to assess whether
turtles are likely to be able to perceive light and what the consequence of the light on their
behaviour is likely to be. The risk assessment should take into account proposed mitigation
and management.
Step 4: Lighting management plan
A lighting management plan for marine turtles should include all relevant project information
(Step 1) and biological information (Step 2). It should outline proposed mitigation. For a range
of specific mitigation measures see the Marine Turtle Light Mitigation Toolbox below. The plan
should also outline the type and schedule for biological and light monitoring to ensure
mitigation is meeting the objectives of the plan and triggers for revisiting the risk assessment
phase of the EIA. The plan should outline contingency options if biological and light monitoring
or compliance audits indicate that mitigation is not meeting the objectives of the plan (e.g. light
is visible on the nesting beach or changes in nesting/hatchling behaviour are observed).
Step 5: Biological and light monitoring and auditing
The success of risk mitigation and light management should be confirmed through monitoring
and compliance auditing. The results should be used to inform continuous improvement.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
60
Relevant biological monitoring is described in Step 2 above. Concurrent light monitoring
should be undertaken and interpreted in the context of how turtles perceive light and within
the limitations of monitoring techniques described in Measuring Biologically Relevant Light.
Auditing as described in the lighting management plan should be undertaken.
Step 6: Review
The EIA should incorporate a continuous improvement review process that allows for
upgraded mitigations, changes to procedures and renewal of the lighting management plan.
Marine Turtle Light Mitigation Toolbox
Appropriate lighting design/lighting controls and light impact mitigation will be site/project and
species specific. Table 6 provides a toolbox of options for use around important turtle habitat.
These options should be implemented in addition to the six Best Practice Light Design
principles. Not all mitigation options will be relevant for every situation. Table 7 provides a
suggested list of light types appropriate for use near turtle nesting beaches and those to avoid.
Two of the most effective approaches for management of light near important nesting beaches
is to ensure there is a tall dark horizon behind the beach such as dunes and/or a natural
vegetation screen and to ensure there is no light on or around the water through which
hatchlings disperse.
Table 6 Light management options specific to marine turtle nesting beaches.
Management Action
Detail
Implement light management actions during the
nesting and hatching season.
Peak nesting season needs to be determined.
Avoid direct light shining onto a nesting beach
or out into the ocean adjacent to a nesting
beach.
Adult turtles nest in lower numbers at lit
beaches (Price et al., 2018).
Maintain a dune and/or vegetation screen
between the nesting habitat and inland sources
of light.
Hatchlings orient towards the ocean by crawling
away from the tall, dark horizon provided by a
dune line and/or vegetation screen.
Maintain a dark zone between turtle nesting
beach and industrial infrastructure.
Avoid installing artificial light within 1.5 km of an
industrial development (Pendoley and
Kamrowski, 2015b).
Install light fixtures as close to the ground as
practicable.
Any new lighting should be installed close to the
ground and reduce the height of existing lights
to the extent practicable to minimise light spill
and skyglow.
Use curfews to manage lighting.
Manage artificial lights using motion sensors
and timers around nesting beaches after dark.
Aim lights downwards and direct them away
from nesting beaches.
Aim light onto the exact surface area requiring
illumination. Use shielding on lights to prevent
light spill into the atmosphere and outside the
footprint of the target area.
Use flashing/intermittent lights instead of fixed
beam.
For example, small red flashing lights can be
used to identify an entrance or delineate a
pathway.
Use motion sensors to turn on lights only when
needed.
For example, motion sensors could be used for
pedestrian areas near a nesting beach.
Prevent indoor lighting reaching beach.
Use fixed window screens or window tinting on
fixed windows, skylights and balconies to
contain light inside buildings.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
61
Management Action
Detail
Limit the number of beach access areas or
construct beach access such that artificial light
is not visible through the access point.
Beach access points often provide a break in
dune or vegetation that protects the beach from
artificial light. By limiting the number of access
points or making the access path wind through
the vegetation, screen light spill can be
mitigated.
Work collectively with surrounding
industry/private land holders to address the
cumulative effect of artificial lights.
Problematic skyglow may not be caused by any
one light owner/manager. By working with other
industry/stakeholders to address light pollution,
the effect of artificial light may be reduced more
effectively.
Manage artificial light at sea, including on
vessels, jetties, marinas and offshore
infrastructure.
Hatchlings are attracted to, and trapped by, light
spill in the water.
Reduce unnecessary lighting at sea.
Extinguish vessel deck lights to minimum
required for human safety and when not
necessary. Restrict lighting at night to navigation
lights only. Use block-out blinds on windows.
Avoid shining light directly onto longlines and/or
illuminating baits in the water.
Light on the water can trap hatchlings or delay
their transit through nearshore waters,
consuming their energy reserves and likely
exposing them to predators.
Avoid lights containing short wavelength
violet/blue light.
Lights rich in blue light can include: metal
halides, fluorescent, halogens, mercury vapour
and most LEDs.
Avoid white LEDs.
Ask suppliers for an LED light with little or no
blue in it or only use LEDs filtered to block the
blue light. This can be checked by examining
the spectral power curve for the luminaire.
Avoid high intensity light of any colour.
Keep light intensity as low as possible in the
vicinity of nesting beaches. Hatchlings can see
all wavelengths of light and will be attracted to
long wavelength amber and red light as well as
the highly visible white and blue light, especially
if there is a large difference between the light
intensity and the ambient dark beach
environment.
Shield gas flares and locate inland and away
from nesting beach.
Manage gas flare light emissions by: reducing
gas flow rates to minimise light emissions;
shielding the flame behind a containment
structure; elevating glow from the shielded flare
more than 30
o
above hatchling field of view;
containing pilot flame for flare within shielding;
and scheduling maintenance activity requiring
flaring outside of turtle hatchling season.
Industrial/port or other facilities requiring
intermittent night-time light for inspections
should keep the site dark and only light specific
areas when required.
Use amber/orange explosion proof LEDs with
smart lighting controls and/or motion sensors.
LEDs have no warmup or cool down limitations
so can remain off until needed and provide
instant light when required for routine nightly
inspections or in the event of an emergency.
Industrial site/plant operators to use head
torches.
Consider providing plant operators with white
head torches (explosion proof torches are
available) for situations where white light is
needed to detect colour correctly or when there
is an emergency evacuation.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
62
Management Action
Detail
Supplement facility perimeter security lighting
with computer monitored infra-red detection
systems.
Perimeter lighting can be operated if night-time
illumination is necessary but remain off at other
times.
No light source should be directly visible from
the beach.
Any light that is directly visible to a person on a
nesting beach will be visible to a nesting turtle or
hatchling and should be modified to prevent it
being seen.
Manage light from remote regional sources (up
to 20 km away).
Consider light sources up to 20 km away from
the nesting beach, assess the relative visibility
and scale of the night sky illuminated by the light
e.g. is a regional city illuminating a large area of
the horizon and what management actions can
be taken locally to reduce the effect i.e. protect
or improve dune systems or plant vegetation
screening in the direction of the light.
Table 7 Where all other mitigation options have been exhausted and there is a human safety
need for artificial light, this table provides commercial luminaire types that are considered
appropriate for use near important marine turtles nesting habitat and those to avoid.
Light type
Suitability for use near marine turtle habitat
Low Pressure Sodium Vapour
High Pressure Sodium Vapour
Filtered* LED
Filtered* metal halide
Filtered* white LED
Narrowband Amber LED
PC Amber
White LED
X
Metal halide
X
White fluorescent
X
Halogen
X
Mercury vapour
X
only if a filter is applied to remove the short wavelength (<500 nm)
light.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
63
Appendix G - Seabirds
Seabirds spend most of their lives at sea, only coming ashore to nest. Many species
are vulnerable to the effects of lighting. Seabirds active at night while migrating,
foraging or returning to colonies are most at risk. Fledglings are more affected by
artificial lighting than adults. Birds can be affected by lights up to 15 km away.
Most common impacts of light on seabirds include disorientation and attraction,
resulting in collisions and/or grounding causing direct or indirect negative effects.
The physical aspects of light that have the greatest impact on seabirds include
intensity and colour (wavelength). Consequently, aside from the reduction of the
spatiotemporal extent of artificial light, management of these aspects of artificial light
will have the most effective result.
Seabirds are birds that are adapted to life in the marine environment (Figure 28). They can be
highly pelagic, coastal, or in some cases spend a part of the year away from the sea entirely.
They feed from the ocean either at or near the sea surface. In general, seabirds live longer,
breed later and have fewer young than other birds and invest a great deal of energy in their
young. Most species nest in colonies, which can vary in size from a few dozen birds to millions.
Many species undertake long annual migrations, crossing the equator or circumnavigating the
Earth in some cases (Ross et al., 1996).
Artificial light can disorient seabirds and potentially cause injury and/or death through collision
with infrastructure on land and at sea. Indirect impacts of artificial lights include increased
predation of grounded birds, collisions with vehicles following grounding, or waterlogging and
drowning following collisions with vessels, and subsequent contamination with chemicals on
board vessels. Particularly high mortality of seabirds occurs through grounding of fledglings
as a result of attraction to lights (Rodríguez et al., 2017c).
Figure 28 Flesh-footed Shearwater (Ardenna carneipes) at sunset. Photo: Richard Freeman.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
64
Conservation Status
Migratory seabird species are protected under international treaties and agreements including
the Convention on the Conservation of Migratory Species of Wild Animals (CMS, Bonn
Convention) and some of the Agreements negotiated under its framework, such as the
Agreement on the Conservation of Albatrosses and Petrels (ACAP) and the Agreement on the
Conservation of African-Eurasian Migratory Waterbirds (AEWA); the Ramsar Convention on
Wetlands; and through the East Asian - Australasian Flyway Partnership (the Flyway
Partnership). Many seabirds are also protected under national environmental legislation.
There are over 350 species of seabirds which are divided into nine orders: Procellariiformes
(albatrosses and petrels); Sphenisciformes (penguins); Gaviiformes (loons); Podicipediformes
(grebes); Anseriformes (waterfowl); Phaethontiformes (tropicbirds); Charadriiformes (gulls,
skuas, skimmers, terns, phalaropes and auks); Pelecaniformes (pelicans); and Suliformes
(frigatebirds, cormorants, gannets and boobies) (Votier and Sherley, 2017).
The IUCN Red List categorises 31% of seabird species as globally threatened (Critically
Endangered, Endangered or Vulnerable) and 11% as Near Threatened (Dias et al., 2019).
Almost half of species (47%) have declining population trends.
Distribution and Habitat
Seabirds breed on all continents of the globe and utilise every sea and ocean on our planet.
Diversity and abundance of seabirds, however, varies spatially and both peak at higher
latitudes, and in the Southern Ocean in particular.
Seabirds spend most of their time at sea but have to return to land to breed. Seabirds are
often vulnerable to predation at their breeding sites and thus, seabirds usually breed on islands
or coastal sand bars, but some species breed far inland in a variety of habitats including
primary rainforest or deserts. Following breeding, seabirds often undertake spectacular
migrations away from their breeding grounds and some travel vast distances across oceans,
sometimes moving between hemispheres, or even circumnavigating the globe. Seabirds can
be affected by artificial light at breeding areas, while foraging and migrating.
For the purposes of these Guidelines, important habitat for seabirds includes those areas
designated as such in wildlife conservation plans and in species specific conservation advice,
for example Key Biodiversity Areas (KBAs).
Effects of Artificial Light on Seabirds
Seabirds have been affected by artificial light sources for centuries. Humans have used fire to
attract seabirds to hunt them for food
and reports of collisions with lighthouses date back to
1880 (Allen, 1880; Murphy, 1936). More recently artificial light associated with the rapid
urbanisation of coastal areas has been linked to increased seabird mortality (Gineste et al.,
2016)
and today, 56 petrel species worldwide are known to be affected by artificial lighting
(Rodríguez et al., 2017ab). Artificial light can disorient seabirds causing collision, entrapment,
stranding, or grounding, and interference with navigation (being drawn off course from usual
migration route) resulting in injury and/or death.
Species active at night are particularly vulnerable as artificial light can disrupt their ability to
orient towards the sea, or even attract birds from sea to land. Additionally, attraction to vessels
due to artificial light at sea can have an impact. Problematic sources of artificial light include

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
65
coastal residential and hotel developments, street lighting, vehicle lights, sporting facility
floodlights, vessel deck and search lights, cruise ships, fishing vessels, gas flares, commercial
squid vessels, security lighting, navigation aids and lighthouses (Ainley et al., 2001; Black,
2005; Raine et al., 2007; Merkel and Johansen, 2011; Rodríguez et al., 2012; Gineste et al.,
2016; Deppe et al., 2017; Rodríguez et al., 2017b; Fischer et al., 2021; Department of
Conservation and Fisheries New Zealand, 2023). Seabirds, particularly petrel species in the
Southern Ocean, can be disoriented by vessel lighting causing collisions and subsequent
injury or death. The effect of artificial light may be exacerbated by moon phase, wind direction
and strength, precipitation, cloud cover and the proximity of nesting sites or migrating sites to
artificial light sources (Troy et al., 2013; Rodríguez et al., 2014; Rodríguez et al., 2015ab;
Deppe et al., 2017; Syposz et al., 2018). The degree of disruption is determined by a
combination of physical, biological and environmental factors including the location, visibility,
colour and intensity of the light, its proximity to other infrastructure, landscape topography,
moon phase, atmospheric and weather conditions and species present.
Seabirds that are active at night while migrating, foraging or returning to colonies and are
directly affected include petrels, shearwaters, albatross, noddies, terns and some penguin
species. Less studied are the effects of light on the colony attendance of nocturnal
Procellariiformes, which could lead to reduced activity, or higher predation risks by avian
predators (Austad et al., 2023). The effects on species that are active during the day, include
extending their activities into the night as artificial light increases perceived daylight hours, are
also little known.
Mechanisms by which light affects seabirds
Many seabird taxonomic groups are diurnal foragers. They rest during dark hours and have
less exposure to artificial light. However, nocturnally active species are more sensitive and
artificial light affects adults and fledglings differently in these species.
Adults are less affected by artificial light than inexperienced younger birds. Many
Procellariiform species (i.e. shearwaters, storm petrels, and petrels) are at risk during their
nocturnal activities. Adult Procellariiformes are vulnerable when returning to and leaving the
nesting colony. They may leave or enter to re-establish their pair bonds with breeding partners,
repair nesting burrows, defend nesting sites or to forage. Adults feed their chick by
regurgitating partially digested food (Imber, 1975). Artificial light disrupts adult nest attendance
and thus affects weight gain in chicks (Cianchetti-Benedetti et al., 2018).
Fledglings are more vulnerable due to the naivety of their first flight, the immature development
of ganglions in the eye at fledging and the potential connection between light and food
(Montevecchi, 2006; Mitkus et al., 2016). Atchoi et al. (2020) proposed that fledglings may be
particularly at risk because of their untrained and undeveloped visual system combined with
their behavioural inexperience. Some fledgling birds do manage to fly over light-polluted areas
and reach the ocean, and it is not clear why some birds are able to do this while others are
grounded (Rodríguez et al., 2022). It may be due to intrinsic factors such as differences in
et al., 2021). Much of the literature
concerning the effect of lighting upon seabirds relates to the synchronised nocturnal mass
exodus of fledglings from their nesting sites (Reed et al., 1985; Le Corre et al., 2002; Raine et
al., 2007; Rodríguez et al., 2015ab; Deppe et al., 2017). For example, fledging
Procellariiformes leave the nesting colony for the sea at night, returning to breed several years
later (Warham, 1990). Emergence during darkness is believed to be a predator-avoidance
strategy
and artificial lighting may make the fledglings more vulnerable to predation (Reed et
al., 1985; Watanuki, 1986). Artificial lights are thought to override the sea-finding cues
provided by the moon and star light at the horizon
and fledglings can be attracted back to
onshore lights after reaching the sea (Telfer et al., 1987; Podolsky et al., 1998; Rodríguez et

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
66
al., 2014). The consequences of exposure to artificial light on the population dynamics and
the overall viability of seabird populations deserve further study (Griesemer and Holmes,
2011).
Eye structure and sensitivities
Seabirds, like most vertebrates, have an eye that is well adapted to see colour. Typically,
diurnal birds have six photoreceptor cells which are sensitive to different regions of the visible
spectrum (Vorobyev, 2003). In all seabirds, their photopic vision (daylight adapted) is most
sensitive in the long wavelength range of the visible spectrum (590 740 nm, orange to red)
while their scotopic (dark adapted) vision is more sensitive to short wavelengths of light (violet
to blue) (Capuska et al., 2011). The eyes of the Black Noddy (Anous minutus) and Wedge-
tailed Shearwaters (Ardenna pacifica), for example, are characterised by a high proportion of
cones sensitive to shorter wavelengths (Hart, 2001). This adaptation is likely due to the need
to see underwater, and the optimum wavelength for vision in clear blue oceanic water is
between 425 and 500 nm. Although many diurnal birds can see in the UV range (less than
380 nm) (Bowmaker et al., 1997), of the over 300 seabird species, only a few have UV
sensitive vision (Capuska et al., 2011). There is no ecological advantage to having many long-
wavelength-sensitive photoreceptors in species foraging in this habitat (Hart, 2001).
Little has been published on vision in penguins. Penguins are visual foragers with the success
of fish capture linked directly to the amount of light present (Cannell and Cullen, 1998). The
eyes of the Humboldt Penguin (Spheniscus humboldti) are adapted to the aquatic
environment, seeing well in the violet to blue to green region of the spectrum, but poorly in the
long wavelengths (red) (Bowmaker and Martin, 1985).
Wavelength, intensity and direction
The intensity of light may be a more important cue than colour for seabirds. Very bright light
will attract them, regardless of colour (Raine et al., 2007). There are numerous, although
sometimes conflicting, reports of the attractiveness of different wavelengths of artificial light to
seabirds. White light has the greatest effect on seabirds as it contains all wavelengths of light
(Wiltschko and Wiltschko, 1999; Rich and Longcore, 2006; Deppe et al., 2017). Seabirds have
reportedly been attracted to the yellow/orange colour of fire (Murphy, 1936), while white
Mercury Vapour and broad-Pterodroma
baraui Puffinus huttoni) than either LPS or HPS Vapour lights
(Deppe et al., 2017). Bright white deck lights and spot lights on fishing vessels attract seabirds
at night, particularly on nights with little moon light or low visibility (Black, 2005; Montevecchi,
2006; Merkel and Johansen, 2011).
A controlled field experiment on short-tailed shearwaters at Phillip Island, Australia tested the
effect of metal halide, LED and HPS lights on fledging groundings (Rodríguez et al., 2017b).
The results suggested that shearwaters were more sensitive to the wider emission spectrum
and higher blue content of metal halide and LED lights than to HPS light. The authors strongly
recommended using HPS, or filtered LED and metal halide lights with purpose designed LED
filtered to remove short wavelength light for use in the vicinity of shearwater colonies.
The first studies of penguins exposed to artificial light at a naturally dark site found they
preferred lit paths over dark paths to reach their nests (Rodríguez et al., 2018). While artificial
light might enhance penguin vision at night, making it easier for them to find their way,
attraction to light could redirect them to undesirable lit areas.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
67
Environmental Impact Assessment of Artificial Light on
Seabirds
As a minimum, infrastructure with artificial lighting that is externally visible should have Best
Practice Lighting Design implemented. Where there is important habitat for seabirds within 20
km of a project, an Environmental Impact Assessment (EIA) should be undertaken. The 20
km buffer for considering important seabird habitat is based on the observed grounding of
seabirds in response to a light source at least 15 km away (Rodríguez et al., 2014). Where
artificial light is likely to affect seabirds, consideration should be given to mitigation measures
at the earliest point in a project development and used to inform the design phase. The spatial
and temporal characteristics of migratory corridors are important for some seabird species.
Species typically use established migratory pathways at predictable times and artificial light
intersecting with an overhead migratory pathway should be assessed in the same way as
ground-based populations. The following sections step through the EIA process with specific
consideration for seabirds.
Associated guidance
• Agreement on the Conservation of Albatrosses and Petrels (ACAP)
Qualified personnel
Lighting design and management and the EIA process should be undertaken by appropriately
qualified personnel. Lighting management plans should be developed and reviewed by
appropriately qualified lighting practitioners who should consult with appropriately trained
marine ornithologists and/or ecologists.
Step 1: Describe the project lighting
The type of information collated during this step should consider the biological Effects of
Artificial Light on Seabirds. Seabirds are susceptible when active at night while migrating,
foraging or returning to colonies. The location and light source (both direct and skyglow) in
relation to breeding and feeding areas should be considered. Seabirds are sensitive to short
wavelengths (blue/violet) but the intensity of lights may be more important than colour.
Step 2: Describe seabird population and behaviour
The species, life stage and behaviour of seabirds in the area and time of interest should be
described. This should include the conservation status of the species; abundance of birds;
regional importance of the population; and seasonality of seabirds utilising the area.
Relevant seabird information can be found in relevant conservation advice; relevant wildlife
conservation plans; scientific literature; and local/Indigenous knowledge.
Where there are insufficient data available to understand the population importance or
demographics, or where it is necessary to document existing seabird behaviour, field surveys
and biological monitoring may be necessary.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
68
Biological monitoring of seabirds
Biological monitoring should be developed, overseen and interpreted by an appropriately
qualified biologist or ornithologist to ensure reliability of the data.
The objectives of monitoring in an area likely to be affected by light include:
• understanding the size and importance of the population
• understanding the habitat use and behaviour of the population (e.g. migrating,
foraging, breeding)
• describing seabird behaviour prior to the introduction/upgrade of light.
The data will be used to inform the EIA process and assess whether mitigation measures are
successful. Suggested minimum monitoring parameters (what is measured) and techniques
(how to measure them) are summarised in Table 8.
Additional seabird monitoring
• Monitor fledging behaviour before a project begins to establish a benchmark for
assessing changes in fledging behaviour during construction and operations.
• Monitor fallout by assessing breeding colonies prior to fledging to assess annual
breeding output/effort and measure against fallout (expecting greater fallout in years
with higher reproductive output).
• Install camera traps at key locations to monitor fallout.
• Conduct nightly assessments of target lighting/areas to identify and collect grounded
birds.
• Conduct observations post-dusk and pre-dawn with night vision goggles to assess
activity/interactions.
• Track movement using land-based radar to determine existing flightpaths (Raine et al.,
2007).
Table 8 Recommended minimum biological information necessary to assess the importance of
a seabird population. Note: the information in this table is not prescriptive and should be
assessed on a case-by-case basis.
Target
Age
Class
Survey Effort
Duration
Reference
Adult
Nesting
In colonial nesting burrow or surface nesting
species with fixed or transient nesting sites, surveys
should be timed to coincide with predicted peak
laying period.
(transects/quadrats) appropriate for nest density to
capture ~100 nests per transect. Status of nests
recorded (used/unused- chick stage).
Transient surface nesting species - estimate of
chicks in crches using aerial or drone footage.
(transects/quadrats) appropriate for nest density to
capture ~100 nests per transect. Status of nests
recorded (used/unused- egg or chick).
Minimum of
two breeding
seasons
Henderson and
Southwood (2016)
Surman and
Nicholson (2014a)

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
69
Fledging
In colonial nesting burrow or surface nesting
species with fixed nesting sites, surveys should be
timed to coincide with predicted max fledging
period.
Minimum of
two breeding
seasons
Henderson and
Southwood (2016)
Surman and
Nicholson (2014b)
As a minimum, qualitative descriptive data on visible light types, location and directivity should
also be collected at the same time as the biological data. Handheld camera images can help
to describe the light. Quantitative data on existing skyglow should also be collected, if possible,
in a biologically meaningful way, recognising the technical difficulties in obtaining these data.
See Measuring Biologically Relevant Light for a review.
Step 3: Risk assessment
The objective is that light should be managed in a way that seabirds are not disrupted within,
or displaced from, important habitat, and they are able to undertake critical behaviours, such
as foraging, reproduction and dispersal. Any disruptions should be considered in the risk
assessment process.
In considering the likely effect of light on seabirds, the assessment should consider the
collected and collated information on the seabirds and the lighting, including the existing light
environment, the proposed lighting design and mitigation/management, and behaviour of
seabirds at the location. Consideration should be given to how the birds perceive light. This
should include both wavelength and intensity information and perspective. To discern
how/whether seabirds are likely to see light, a site visit should be made at night. Similarly,
consideration should be given to how seabirds will see light when in flight.
Using this perspective, the type and number of lights should be considered/modelled to
determine whether seabirds are likely to perceive light and what the consequence of the light
on their behaviour is likely to be.
Step 4: Lighting management plan
A lighting management plan should include all relevant project information (Step 1) and
biological information (Step 2). It should outline proposed mitigation. For a range of seabird
specific mitigation measures please see the Seabird Light Mitigation Toolbox below. The plan
should also outline the type and schedule for biological and light monitoring to ensure
mitigation is meeting the objectives of the plan and triggers for revisiting the risk assessment
phase of the EIA. The plan should outline contingency options if biological and light monitoring
or compliance audits indicate that mitigation is not meeting objectives (e.g. light is mitigated
appropriately and impacts are not reduced).
Step 5: Biological and light monitoring and auditing
The success of the impact mitigation and light management should be confirmed through
monitoring and compliance auditing and the results used to facilitate an adaptive management
approach for continuous improvement.
Relevant biological monitoring is described in Step 2 above. Concurrent light monitoring
should be undertaken and interpreted in the context of how seabirds perceive light and within
the limitations of monitoring techniques described in Measuring Biologically Relevant Light.
Auditing, as described in the lighting management plan, should be undertaken.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
70
Step 6: Review
The EIA should incorporate a continuous improvement review process that allows for
upgraded mitigations, changes to procedures and renewal of the lighting management plan.
Seabird Light Mitigation Toolbox
Appropriate lighting design/lighting controls and mitigating the effect of light will be site/project
and species specific. Table 9 provides a toolbox of management options relevant to seabirds.
These options should be implemented in addition to the six Best Practice Light Design
principles. Not all mitigation options will be practicable for every project. Table 10 provides a
suggested list of light types appropriate for use near important seabird habitat and those to
avoid.
A comprehensive review of the effect of land based artificial lights on seabirds and mitigation
techniques found the most effective measures were:
• turning lights off, particularly during the fledgling periods
• removing external lights and closing window blinds to shield internal lights
• shielding light sources and preventing upward light spill
• modification of light wavelengths
• reducing traffic speed limits and display of warning signs
• implementing a rescue programme for grounded birds (Rodríguez et al., 2017c)
• keeping light intensity as low as possible. Most bird groundings are observed in very
brightly lit areas (Rodríguez et al., 2017c).
Table 9 Light management options for seabirds.
Management Action
Detail
Maintain a dark zone around
important seabird habitat.
Avoid installing lights or manage all outdoor lighting
within 3 km of important seabird habitat (recorded
median distance between nests and grounding
location) (Rodríguez et al., 2015b).
Turn off lights during fledgling
season.
If not possible to extinguish lights completely, consider
dimming options, or changes on light spectra
(preferably to reduce blue emissions). New moon
periods and when conditions are rainy or foggy are
high risk periods and when mitigation efforts should be
increased.
Use curfews to manage lighting.
Extinguish lights around seabird breeding habitat
during the fledgling period by dusk as fledglings leave
their nest early in the evening.
Aim lights downwards and direct
them away from nesting areas.
Aim light onto only the surface area requiring
illumination. Use shielding to prevent light spill into the
atmosphere and outside the footprint of the target area.
Use motion sensors to turn lights on
only when needed.
Use motion sensors for pedestrian or street lighting
within at least 3 km of important seabird habitat,
although effects may extend further and the latest
research should be consulted when determining
distances.
Avoid high intensity light of any
colour.
Keep light intensity as low as possible in the vicinity of
important seabird habitat.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
71
Use luminaires with spectral content
appropriate for the species present.
Consideration should be given to avoid specific
wavelengths that are problematic for the species of
interest. In general, this would include avoiding lights
rich in blue light.
Prevent indoor lighting reaching
outdoor environment.
Use fixed window screens or window tinting on fixed
windows and skylights to contain light inside buildings.
Manage artificial light on jetties,
wharves, marinas, etc.
Fledglings and adults may be attracted to lights on
marine facilities and become grounded or collide with
infrastructure.
Reduce unnecessary outdoor, deck
lighting on all vessels and
permanent and floating oil and gas
installations in known seabird
foraging areas at sea.
Extinguishing outdoor/deck lights when not necessary
for human safety and restrict lighting at night to
navigation lights. Use block-out blinds on all portholes
and windows.
Night fishing should only occur with
minimum deck lighting.
Avoid shining light directly onto
fishing gear including longlines in
the water.
Ensure lighting enables recording of
any incidental catch, including by
electronic monitoring systems.
Light on the water at night can attract seabirds to
deployed fishing gear increasing the risk of seabird
bycatch (i.e. killing or injuring birds).
Minimum deck lighting should not breach minimum
standards for safety and navigation.
Record vessel strikes and bycatch and report these
data to regulatory authorities.
Vessels working in seabird foraging
areas during breeding season
should implement a seabird
management plan to prevent vessel
strikes.
Lights at sea should be managed similarly to lights on
land to avoid vessel strikes (collisions with or
unintentional landings on vessel and associated
superstructure) and impacts (direct or indirect) thereof.
For example, see Department of Conservation and
Fisheries New Zealand, 2023 and Managing artificial
lights to reduce seabird vessel strikes
Use flashing/intermittent lights
instead of fixed beam.
For example, small red flashing lights can be used to
identify an entrance or delineate a pathway.
Shield gas flares and locate inland
and away from important seabird
habitat.
Manage gas flare light emissions by: reducing gas flow
rates to minimise light emissions; shielding the flame
behind a containment structure; containing the pilot
flame for flare within shielding; and scheduling
maintenance activity requiring flaring outside of the
seabird breeding season or during the day.
Minimise flaring on offshore oil and
gas production facilities.
Consider reinjecting excess gas instead of flaring,
particularly on installations on migratory pathways.
In facilities requiring intermittent
night-time inspections, turn on lights
only during the time operators are
moving around the facility.
Use appropriate wavelength lights with smart lighting
controls. LEDs have no warmup or cool down
limitations so can remain off until needed and provide
instant light when required for routine nightly
inspections or in the event of an emergency.
Ensure industrial site/plant
operators use head torches.
Consider providing operators with white head torches
where appropriate for situations where white light is
needed to detect colour correctly or in an emergency.
Supplement facility perimeter
security lighting with computer
Perimeter lighting can be operated when night-time
illumination is necessary but otherwise remain off.
UNEP/CMS/COP14/Doc.30.4.4/Annex 3
72
monitored infrared detection
systems.
Tourism operations around seabird
colonies should manage torch
usage.
Consideration should be given to educational signage
around seabird colonies where tourism visitation is
generally unsupervised.
Design and implement a rescue
programme for grounded birds.
This will not prevent birds grounding, but it is an
important management action in the absence of
appropriate light design. Rescue programmes have
proven useful to reducing mortality of seabirds. The
programme should include documentation and
reporting of data about the number and location of
rescued birds to regulatory authorities. Ensure birds
are released in a safe area and at appropriate time to
avoid predators.
Table 10 Where all other mitigation options have been exhausted and there is a human safety
need for artificial light, this table provides commercial luminaires recommended for use near
seabird habitat and those to avoid.
Light type
Suitability for use near
migratory seabird habitat
Low Pressure Sodium Vapour
High Pressure Sodium Vapour
Filtered* LED
Filtered* metal halide
Filtered* white LED
LED with appropriate spectral properties for species present
White LED
Metal halide
White fluorescent
Halogen
Mercury vapour
only if a filter is applied to remove the problematic
wavelength light.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
73
Appendix H - Migratory Shorebirds
There is evidence that night-time lighting of migratory shorebird foraging areas may
benefit the birds by allowing greater visual foraging opportunities. However, where
nocturnal roosts are artificially illuminated, shorebirds may be displaced, potentially
reducing their local abundance if the energetic cost to travel between suitable
nocturnal roosts and foraging sites is too great.
Artificial lighting could also act as an ecological trap by drawing migratory
shorebirds to foraging areas with increased predation risk. Overall, the effect of
artificial light on migratory shorebirds remains understudied and consequently any
assessment should adopt the precautionary principle and manage potential negative
effects from light unless demonstrated otherwise.
Shorebirds, also known as waders, inhabit the shorelines of coasts and inland water bodies
for most of their lives. They belong to the order Charadriiformes. Most are from two taxonomic
families, the Sandpipers (Scolopacidae) and the Plovers (Charadriidae). They are generally
distinguished by their relatively long legs, often long bills, and most importantly, their
associations with wetlands at some stages of their annual cycles (van de Kam et al., 2004).
At least 215 shorebird species have been described
and their characteristics include long
lifespans but low reproductive output (Colwell, 2010). Many species have specialised bills for
feeding on different prey in wetlands. The bills of many species contain sensory organs to
detect the vibrations of prey inside the substrate. Shorebirds are often gregarious during the
non-breeding season, which is probably a mechanism to reduce individual predation risk and
increase the chance of locating profitable feeding patches (Cresswell, 1994; Piersma and
Baker, 2000). Over 60 per cent of shorebird species migrate. Some are transoceanic and
transcontinental long-distance migrants capable of flying for many days non-stop. Bar-tailed
godwits (Limosa lapponica), for example, have been recorded flying non-stop for up to 11,500
km (Battley et al., 2012).
Figure 29 Curlew Sandpipers (Calidris ferruginea). Photo: Brian Furby.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
74
Conservation Status
Migratory shorebird species are protected under international treaties and agreements
including the Convention on the Conservation of Migratory Species of Wild Animals (CMS,
Bonn Convention), the Ramsar Convention on Wetlands, and the East Asian - Australasian
Flyway Partnership, the Agreement on the Conservation of African-Eurasian Migratory
Waterbirds (AEWA) and Americas partnership. Many species are also protected under
national environmental legislation.
Forty-one per cent of populations covered by AEWA are decreasing in the short-term, 29%
are stable and 30% are increasing (UNEP/AEWA Secretariat, 2021). Long-term trends are
similar (43%, 23% and 34% respectively). The proportion of decreasing populations is
particularly high in the Central and Southwest Asian, Eastern and Southern African and Sub-
Saharan African flyways. In various parts of the flyways, large-scale industrial environments
and reclamations threaten migratory species by removing the main stop-over habitat. Trend
data are available for 35 shorebird populations using the Western Atlantic Flyway with 65% of
those populations declining (Watts et al., 2015). Piersma et al. (2016) reported that habitat
loss along the Yellow Sea is contributing to declining shorebird numbers along the East Asian-
Australasian Flyway.
Some regions have increasing populations. The proportion of increasing populations is
particularly high in Western and Central Africa, the Atlantic part of the Palearctic and in the
Black Sea and Mediterranean, Sahelian and East Atlantic flyways (UNEP/AEWA Secretariat,
2021).
Distribution and Habitat
Migratory shorebirds are found in almost all countries, with some present throughout the year
in most. Peak abundance occurs in Spring/Summer in countries where they breed. In breeding
areas many species use inland habitats, in particular tundra but also various types of wetlands.
In non-breeding areas they are predominantly associated with coastal wetland habitats
including estuaries and intertidal wetlands, beaches, saltmarsh, mangrove fringes, wet
grasslands, ephemeral freshwater and salt lakes, pastures, rice paddies, tilled land, sewage
treatment plants, irrigation canals, sports fields and golf courses.
Migratory shorebirds use flyways during their migrations. For a detailed review of bird flyways
see UNE
a single migratory species, a group of migratory species or a distinct population of a given
migratory species completes all components of its annual cycle (breeding, moulting, staging,
non-breeding etc.). For some species and groups of species these flyways are distinct
The East Asian-Australasian Flyway, for example, stretches from the Russian Far East and
Alaska, through East Asia and South-east Asia, to Australia and New Zealand with 397
internationally recognised sites considered important for migratory shorebirds along it
(Bamford et al., 2008). It is home to over 50 million migratory waterbirds from over 250 different
populations (EAAFP, 2018).
Similarly, the Central Asian Flyway (CAF) covers a large continental area of Eurasia between
the Arctic and Indian Oceans and the associated island chains (CMS, 2023b). The CAF
comprises several important migration routes of waterbirds, most of which extend from the

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
75
northernmost breeding grounds in the Russian Federation (Siberia) to the southernmost non-
breeding (wintering) grounds in West and South Asia, the Maldives and the British Indian
Ocean Territory. The CAF covers at least 279 populations of 182 migratory waterbird species,
including 29 globally threatened and near-threatened species, which breed, migrate and
winter within the region.
Important habitat for migratory shorebirds
For the purposes of these Guidelines, important habitat for migratory shorebirds includes all
areas that are recognised, or eligible for recognition as nationally or internationally important
habitat.
• Internationally important habitat are those wetlands that regularly support one per
cent of the individuals in a population of one species or subspecies; or a total
abundance of at least 20,000 or more waterbirds (Ramsar Sites Criteria).
• Nationally important habitat may vary according to country. For example, in Australia
nationally important habitat are those wetlands that support 0.1 per cent of the flyway
population of a single species; 2000 migratory shorebirds; or 15 migratory shorebird
species.
East Asian-Australasian Flyway
Many of the northern hemisphere breeders nest in the arctic or sub-arctic tundra during the
boreal summer (May July) and spend the non-breeding season (August April) in Australia
or New Zealand. They usually spend five to six months on the non-breeding grounds, where
they complete their basic (non-breeding plumage) moult, and later commence a pre-alternate
(breeding plumage) moult prior to their northward migration. While undergoing their pre-
alternate moult, shorebirds also consume an increased amount of prey to increase their fat
storages, permitting them to travel greater distances between refuelling sites. Shorebirds
refuel in East Asia during their northward migration, but during southward migration, some
individuals travel across the Pacific, briefly stopping on islands to refuel. Shorebirds migrating
across the Pacific typically have non-breeding grounds in Eastern Australia and New Zealand.
Shorebirds returning to non-breeding grounds in Western and Northern Australia, once again
pass through East Asia on their southward journey.
Western Atlantic Flyway
Many northern hemisphere breeders nest in the arctic or sub-arctic tundra during the boreal
summer (May July), though other species are common in the grasslands of western and
central North America, and still others are common in coastal wetlands. Most populations of
many species spend the non-breeding season (August April) en route to or in more southern
locations, including significantly far south into the southern hemisphere. Birds usually spend
five to six months on the non-breeding grounds, regardless of their distance from the breeding
areas, where they complete their basic (non-breeding plumage) moult, and later commence a
pre-alternate (breeding plumage) moult prior to their northward migration. While undergoing
their pre-alternate moult, shorebirds also consume an increased amount of prey to increase
their fat storages, permitting them to travel greater distances between refuelling sites.
Shorebirds refuel in portions of northern South America, but especially in southern and eastern
North America during their northward migration; during southward migration, some individuals
travel over land through the central regions of the continent, though also across the eastern
Pacific and western North Atlantic, depending on the species and the population, stopping to
refuel only in unfavourable weather conditions. A common feature for many birds is their
reliance on inland or coastal wetland habitats at some stages in their annual life-histories. In
many migratory shorebirds, despite the vast distances they cover every year, they spend most

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
76
of their time on coastal wetlands except for the two months of nesting when they use the
tundra or taiga habitats. However, productive coastal wetland is localised, which means large
proportions, or even entire populations, gather at a single site during stopover or non-breeding
season. Delaware Bay, for example, is the most important spring stopover area for the North
American population of red knot (Calidris canutus rufa) with up to 90% of the population
stopping there within a very narrow time window (American Bird Conservancy, 2023). For red
knots (C.c. rogersi and C.c. piersmai) migrating north along the East Asian-Australasian
Flyway at least 45% and perhaps close to 100% stopover in Bohai Bay, primarily at the Nanpu
tidal flat in China (Mu et al., 2022). Wetlands commonly used include coastal mudflats and
sandflats, sandy beaches, saltmarsh and mangrove fringes, ephemeral freshwater wetlands
and damp grasslands.
The coastal intertidal wetlands favoured by many migratory shorebirds are a dynamic
ecosystem strongly influenced by the tidal cycle. This is part of the critical transition zones
between land, freshwater habitats, and the sea. Throughout migration flyways intertidal
wetlands have been susceptible to heavy modification for the development of farmlands,
aquaculture, salt mining, ports and industry.
The daily activity pattern of shorebirds at coastal wetlands is not only determined by daylight,
but also tidal cycle (Colwell, 2010). They feed on the exposed tidal wetland during low tide
and roost during high tide as their feeding areas are inundated. The birds feed during both the
day and night, especially in the lead-up to migration (Lourenço et al., 2008; Santiago-Quesada
et al., 2014).
Roost site selection can vary between day and night. Shorebirds often use diurnal roosts
nearest to the intertidal feeding area and may travel further to use safer nocturnal roosts but
at greater energetic cost (Dias et al., 2006; Rogers et al., 2006b). Roosting habitat can also
vary between day and night. For example, the dunlin (Calidris alpina), in California, had a
greater use of pasture at night (which tended to be less affected by artificial light and
disturbances) and relied less on their diurnal roosts of islands and artificial structures such as
riprap and water pipes (Conklin and Colwell, 2007).
Foraging behaviours differ between day and night, and between seasons (Lourenço et al.,
2008; McNeil et al., 1993). Shorebirds typically show a preference for daytime foraging, which
occurs over a greater area, and at a faster rate, than nocturnal foraging (Lourenço et al., 2008).
Increased prey availability, avoidance of daytime predation and disturbance are some reasons
for nocturnal foraging (McNeil et al., 1993). Two basic types of foraging strategies have been
described: visual and tactile (touch-based) foraging, with some species switching between
these strategies. Tactile feeders such as sandpipers can use sensory organs in their bills to
detect prey inside the substrate in the dark and can switch to visual foraging strategy during
moonlit nights to take advantage of the moonlight (McNeil et al., 1993). Visual feeders such
as plovers, have high densities of photo receptors, especially the dark-adapted rods, which
allow foraging under low light conditions (McNeil et al., 1993; Rojas et al., 1999). Plovers have
been shown to employ a visual foraging strategy during both the day and night, whereas
sandpipers can shift from visual foraging during the day, to tactile foraging at night, likely due
to less efficient night vision (Lourenço et al., 2008).
Effects of Artificial Light on Migratory Shorebirds
Artificial light can disorient flying birds, affect stopover selection, and cause their death through
collision with infrastructure (McLaren et al., 2018). Birds may starve as a result of disruption
to foraging, hampering their ability to prepare for breeding or migration.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
77
Vision in migratory shorebirds
There is a dearth of literature on light perception in migratory shorebirds with most studies
their response to different wavelengths of light.
Birds in general are known to be attracted to, and disoriented by, artificial lights. This could be
a result of being blinded by the intensity of light that bleaches visual pigments and therefore
failing to see visual details (Verheijen, 1985)
or interference with the magnetic compass used
by the birds during migration (Poot et al., 2008). An attraction to conventional artificial night
lightings may lead to other adverse consequences such as reducing fuel stores, delaying
migration, increasing the chance of collision and thereby, injury and death (Gauthreaux and
Belser, 2006).
Biological impacts on migratory shorebirds
Artificial lighting has been shown to influence the nocturnal foraging behaviour in shorebirds
(Santos et al., 2010; Dwyer et al., 2013). Santos et al. (2010) demonstrated that three species
of plover (common ringed plover Charadrius hiaticula, Kentish plover Charadrius alexandrina
and grey plover Pluvialis squatarola) and two species of sandpiper (dunlin and common
redshank Tringa totantus) had improved foraging success by exploiting sites where
streetlights provided extra illumination. Similarly, Dwyer et al. (2013) showed artificial light
generated from a large industrial site significantly altered the foraging strategy of common
redshanks within an estuary. The greater nocturnal illumination of the estuary from the
industrial site allowed the birds to forage for extended periods using a visual foraging strategy,
which was deemed a more effective foraging behaviour when compared to tactile foraging
(Dwyer et al., 2013). However, patterns of shifts to nocturnality were species-specific and
increased foraging success should not necessarily be considered a net benefit for any species.
For shorebirds increased foraging success may decrease their food sources and negatively
impact them long-term. Owens et al. (2020) document light pollution as contributing to insect
declines and that light pollution could be disrupting whole ecosystems and, therefore, cannot,
at this stage, be considered to benefit shorebirds.
Although shorebirds may be attracted to foraging areas with greater nocturnal illumination,
artificial light near nocturnal roosting sites may displace the birds. Rogers et al. (2006a)
studied the nocturnal roosting habits of shorebirds in north-western Australia, and suggested
nocturnal roost sites with low exposure to artificial lighting (e.g. streetlights and traffic) were
selected, and where the risk of predation was perceived to be low. The study also found
nocturnal roosts spatially differed from diurnal roosts and required increased energetic cost to
access as the distance between nocturnal roosts and foraging areas was greater than the
distance between diurnal roost sites and the same foraging areas (Rogers et al., 2006b). The
overall density of shorebirds in suitable foraging areas is expected to decline with increased
distance to the nearest roost, due to the greater energetic cost travelling between areas (Dias
et al., 2006; Rogers et al., 2006b). The artificial illumination (or lack thereof) of nocturnal roost
sites is therefore likely to significantly influence the abundance of shorebirds in nearby foraging
areas. Intermittent or flashing lights could flush out the shorebirds and force them to leave the
area, especially if the light is persistent (Choi pers. obs. 2018, Straw pers. comm. 2018).
Artificial light can affect birds in flight. Not only can bright light attract airborne migrants, but
artificial light can also affect stop-over selection in long distance migrators which can impact
on successful migration and decrease fitness (Longcore et al., 2013; McLaren et al., 2018).
Similarly, Roncini et al. (2015) reported on interactions between offshore oil and gas platforms
and birds in the North Sea and found these were likely to include migratory shorebirds. Impacts
are likely to be region, species and platform specific.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
78
Environmental Impact Assessment of Artificial Light on
Migratory Shorebirds
As a minimum, Best Practice Lighting Design should be implemented on infrastructure with
externally visible artificial lighting. Where there is important habitat for migratory shorebirds
within 20 km of a project, consideration should be given as to whether that light is likely to
have an effect on those birds. The following sections step through the framework for managing
artificial light, with specific consideration for migratory shorebirds. The 20 km buffer is based
on a precautionary approach that skyglow can cause a change in behaviour in other species
up to 15 km away (Rodríguez et al., 2014).
Where artificial light is likely to affect migratory shorebirds, consideration should be given to
mitigation measures at the earliest point in a project and used to inform the design phase.
It is important to recognise the spatial and temporal characteristics of migratory corridors for
some migratory shorebird species. Species typically use established migratory pathways at
predictable times and artificial light intersecting with an overhead migratory pathway should
be assessed in the same way as for ground-based populations.
Associated guidance
• AEWA Plan of Action for Africa 2019-2027
• East Asian – Australasian Flyway Partnership 2019-2028 Strategic Plan
• Central Asian Flyway Action Plan to Conserve Migratory Waterbirds and their Habitats
• Approved conservation advice
Qualified personnel
Lighting design/management and the EIA process should be undertaken by appropriately
qualified personnel. Plans should be developed and reviewed by appropriately qualified
lighting practitioners who should consult with appropriately trained marine ornithologists or
ecologists.
Step 1: Describe the project lighting
The information collated during this step should consider the biological effects of artificial light
on migratory shorebirds. They can be affected by light when foraging or migrating at night.
Artificial light at night may also affect their selection of roost site. The location and light source
(both direct and skyglow) in relation to feeding and resting areas should be considered,
depending on whether the birds are active or resting at night. Shorebirds are sensitive to short
wavelength (blue/violet) light with some species able to detect UV light. However, the intensity
of lights may be more important than colour.
Step 2: Describe the migratory shorebird population and behaviour
The species, and behaviour of shorebirds in the area of interest should be described. This
should include the conservation status of the species; abundance of birds; how
widespread/localised is the population; the migratory corridor location and timing or usage;
the regional importance of the population; the number of birds in the area in different seasons;
and their night-time behaviour (resting or foraging).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
79
Relevant shorebird information should be sought from the scientific literature, local/Indigenous
knowledge and other relevant sources for the location.
Where there is insufficient data to understand the population importance or demographics, or
where it is necessary to document existing shorebird behaviour, field surveys and biological
monitoring may be necessary.
Biological monitoring of migratory shorebirds
Monitoring associated with a project should be developed, overseen and results interpreted
by appropriately qualified biologists to ensure reliability of the data. The objective is to collect
data on the abundance of birds and their normal behaviour. The data will be used to inform
the EIA and assess whether mitigation measures are successful. Suggested minimum
monitoring parameters (what is measured) and techniques (how to measure them) are
summarised in Table 11.
Table 11 Recommended minimum biological information necessary to assess the importance
of a migratory shorebird population. Note: the information in this table is not prescriptive and
should be assessed on a case-by-case basis.
Survey Effort
Duration
Reference
Timing of surveys will depend on
seasonal patterns in site use by
shorebirds and the functions
(breeding, stop-over, wintering)
the site is used for.
For non-breeding birds, multi-
species surveys are typically
carried out in January (for
northern hemisphere breeding
species) and July (for certain
Afrotropical breeding species).
For breeding birds, the best time
to survey will depend on both the
timing of the breeding season of
the species concerned and the
precise period within the breeding
season at which it is most
effective to conduct a survey.
During migration periods, the
exact timing of spring or autumn
surveys will depend on the
phenology of the species
concerned.
Two hours
before and after
predicted high
tide.
AEWA Guidelines on Waterbird
Monitoring
Monitoring migratory shorebird populations
• Monitor the population (during different seasons) to establish a benchmark for
assessing abundance before, during and after construction, and during operations to
detect project-related change.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
80
• Quantify the diurnal and nocturnal habitat use and movement in relation to tidal cycle
(both high and low tides during the neap and spring tide cycles) in the area under
baseline conditions to compare with light-affected conditions during construction and
operations.
• Measure nocturnal light levels at foraging sites and nocturnal roost sites before and
after the construction period of a project.
• Monitor nocturnal roost sites using acoustic recording devices and/or infrared cameras
to determine nocturnal roost site use following the introduction of artificial light.
As a minimum, qualitative descriptive data on visible light types, location and directivity
should also be collected at the same time as the biological data. Handheld camera images
can help to describe the light. Quantitative data on existing skyglow should be collected, if
possible, in a biologically meaningful way, recognising the technical difficulties in obtaining
these data. See Measuring Biologically Relevant Light for a review.
Step 3: Risk assessment
The objective of these Guidelines is that light should be managed so that shorebirds are not
disrupted within or displaced from important habitat and are able to undertake critical
behaviours such as foraging, roosting and dispersal. These consequences should be
considered in the risk assessment process. At important shorebird habitats, roosting and
foraging numbers should remain constant and foraging birds should not be startled or at
increased risk from predators as a result of increased illumination.
The assessment should consider the existing light environment, the proposed lighting design
and mitigation/management, the behaviour of shorebirds at the location, and how the birds
perceive light. This should include wavelength and intensity information and perspective. To
understand how/whether shorebirds are likely to see light, a site visit should be made at night
and the area viewed from the intertidal flats and roosting areas. Similarly, consideration should
be given to how shorebirds will see light when in flight and along flyways during migration
periods.
The type and number of artificial lights should then be considered to assess whether the birds
are likely to perceive the light, and the possible consequences of light on their behaviour.
Step 4: Lighting management plan
This plan should include all relevant project information (Step 1) and biological information
(Step 2). It should outline proposed mitigation. For a range of shorebird specific mitigation
measures see the Migratory Shorebird Light Mitigation Toolbox below. The plan should also
outline the type and schedule for biological and light monitoring to ensure mitigation is meeting
the objectives of the plan and triggers for revisiting the risk assessment phase of the EIA. The
plan should outline contingency options if biological and light monitoring or compliance audits
indicate that mitigation is not meeting the objectives of the plan (e.g. light is visible on intertidal
flats, shorebirds cease using resting areas, or birds are grounding or colliding with fixed or
floating infrastructure, or migrating birds cease using a migratory corridor).
Step 5: Biological and light monitoring and auditing
The success of the plan should be confirmed through monitoring and compliance auditing.
The results should be used to facilitate an adaptive management approach for continuous
improvement.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
81
Biological monitoring is described in Step 2 above. Concurrent light monitoring should be
undertaken and interpreted in the context of how the birds perceive light and within the
limitations of monitoring techniques described in Measuring Biologically Relevant Light.
Auditing, as described in the plan, should be undertaken.
Step 6: Review
The EIA should incorporate a continuous improvement review process that allows for
upgraded mitigations, changes to procedures and renewal of the lighting management plan.
Migratory Shorebird Light Mitigation Toolbox
All projects should incorporate the Best Practice Light Design Principles. Appropriate lighting
controls and light impact mitigation will be site/project and species specific. Table 12 provides
a toolbox of options that could be implemented in addition to the six Best Practice Light Design
principles. Not all mitigation options will be relevant for all situations. Table 13 provides a
suggested list of light types appropriate for use near rookeries or roosting sites and those to
avoid.
Table 12 Light management actions specific to migratory shorebirds.
Management Action
Detail
Implement actions when birds are likely
to be present. This includes peak
migration periods (flyway locations).
Migration periods need to be identified. Data from
citizen science could be used to identify annual-
cycle phenology, for example eBird.
No light source should be directly visible
from foraging or nocturnal roost habitats,
or from migratory pathways.
Any light that is directly visible to a person
standing in foraging or nocturnal roost habitats will
potentially be visible to a shorebird and should be
modified to prevent it being seen. Similarly, lights
should be shielded such that they are not visible
from the sky.
Do not install fixed light sources in
nocturnal foraging or roost areas.
Installing light sources (e.g. light poles) within
shorebird habitat may permanently reduce the
available area for foraging or roosting and provide
vantage points for predators (e.g. raptors) during
the day.
Prevent mobile light sources shining into
nocturnal foraging and roost habitat.
The light from mobile sources such as mobile
lighting towers, head torches or vehicle headlights
should be prevented from aiming into nocturnal
foraging or roost areas, as this can cause
immediate disturbance.
Maintain a natural barrier (e.g. dune
and/or vegetation screen) between
nocturnal foraging and roost areas, and
sources of artificial light.
Reducing the exposure of shorebirds to artificial
light will reduce the risk of predation and
disturbance.
Maintain a dark zone between nocturnal
foraging and roost habitats and sources
of artificial lights.
Creating a dark zone between artificial lights and
shorebird habitat will reduce disturbances to
shorebirds.
Use curfews to manage lighting near
nocturnal foraging and roosting areas in
coastal habitats. For example, manage
Curfews should also consider the tidal cycle if the
artificial lighting is located coastally, e.g. extinguish
lighting from two hours before high tide, until two

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
82
artificial lights using motion sensors and
timers from dusk until dawn.
hours after high tide, while shorebirds are
potentially roosting.
Use of flashing/intermittent lights instead
of fixed beam.
For example, small red flashing lights can be used
to identify an entrance or delineate a pathway. The
timing of when lights flash must follow a
predictable, well-spaced pattern.
Use motion sensors to turn lights on
only when needed.
For example, installing motion-activated pedestrian
lighting within 500 m of nocturnal foraging or roost
areas may reduce the amount of time the habitat is
exposed to artificial light.
Manage artificial light on jetties and
marinas.
Shorebirds will often roost on breakwaters and
jetties, so allowing dark areas in such places may
provide a safe area for shorebirds to roost.
Reduce deck lighting to minimum
required for human safety on vessels
moored near nocturnal foraging and
roost areas, and those operating
offshore.
Extinguish deck lights when not necessary and
restrict lighting at night to navigation lights only.
Offshore vessels should direct light inwards,
particularly during the migration periods when
shorebirds are potentially overhead.
Record bird strike or incidental capture and report
these interactions to regulatory authorities.
Minimise night-time flaring on offshore
oil and gas production facilities.
Consider reinjecting excess gas instead of flaring.
Schedule maintenance flaring during daylight
hours.
Record bird strike or incidental capture and report
these interactions to regulatory authorities.
Use luminaires with spectral content
appropriate for the species present.
Consideration should be given to avoid specific
wavelengths that are problematic for the species of
interest. In general this would include avoiding
lights rich in blue light, however, some species are
sensitive to yellow light and other mitigation may
be required.
Avoid high intensity light of any colour.
Keeping light intensity as low as possible in the
vicinity of nocturnal foraging and roost areas will
minimise impact.
Prevent indoor lighting reaching
migratory shorebird habitat.
Use fixed window screens or window tinting on
fixed windows and skylights to contain light inside
buildings.
In facilities requiring intermittent night
inspections, turn lights on only during
the time operators are moving around
the facility.
Use appropriate wavelength, explosion proof LEDs
with smart lighting controls and/or motion sensors.
LEDs have no warmup or cool down limitations so
can remain off until needed and provide instant
light when required for routine nightly inspections
or in the event of an emergency.
Industrial site/plant operators to use
personal head torches.
Consider providing plant operators with white head
torches (explosion proof torches are available) for
situations where white light is needed to detect
colour correctly, or in the event of an emergency.
Operators should avoid shining light across
nocturnal foraging or roost areas as this can cause
disturbance.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
83
Supplement facility perimeter security
lighting with computer monitored
infrared detection systems.
Perimeter lighting can be operated when night-
time illumination is necessary but remain off at
other times.
Table 13 Where all other mitigation options have been exhausted and there is a human safety
need for artificial light, the following table provides commercial luminaires recommended for
use near migratory shorebird habitat and those to avoid.
Light type
Suitability for use near
migratory shorebird habitat
Low Pressure Sodium Vapour
High Pressure Sodium Vapour
Filtered* LED
Filtered* metal halide
Filtered* white LED
LED with appropriate spectral properties for species
present
White LED
Metal halide
White fluorescent
Halogen
Mercury vapour
only if a filter is applied to remove the problematic
wavelength light.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
84
Appendix I - Migratory Landbirds
Light pollution impacts migratory landbirds at breeding and overwintering sites but
the period of birds’ annual cycles when they are migrating between the two and
associated with stopover habitats while birds are in transit may represent the most
serious times of concern. Collision is a serious threat and can take place when
nocturnally migrating landbirds are attracted to and disoriented by lights of buildings
or other structures. Such collisions may occur directly, while migrating at night, or
indirectly, when they crash into reflective surfaces on mornings following attraction
to built environment areas after a night’s migration.
Other threats from light pollution include physiological and behavioural impacts that
alter aspects of annual, diel and circadian ecology and phenology. Reducing artificial
light emission into the environment during periods of intense migration can reduce
the negative impacts on migratory landbirds. Weather forecasts and radar can be
used to predict these intense migration periods when mitigation is most important.
This appendix covers migratory landbirds although much of the information included is also
relevant for mitigating against the impacts of light pollution on non-migratory landbirds. There
African-
Eurasian Migratory Landbirds Action Plan (AEMLAP)
Gruiformes, Charadriiformes, Columbiformes, Caprimulgiformes, Apodiformes, Cuculiformes,
Coraciiformes, Piciformes and Passeriformes, which are principally ecologically dependent on
terrestrial habitats and for which the entire population, or significant proportions of the
population, cyclically and predictably cross one or more national juris
Not all species/populations of the orders listed are covered by AEMLAP are regarded as
landbirds, however, and, indeed, some Charadriiformes are covered by Appendix H -
Migratory Shorebirds (families Glareolidae, Scolopacidae and Charadriidae). The
Memorandum of Understanding on the Conservation of Migratory Birds of Prey in Africa and
Eurasia covers Falconiformes and Strigiformes.
BirdLife International Data
Zone gives a result of 1,290 extant species including (amongst many others) species in the
families Tyrannidae (Tyrant-flycatchers) (113 species), Accipitridae (Hawks, Eagles) (80),
Muscicapidae (Old World Flycatchers and Chats) (76), Hirundinidae (Swallows and martins)
(58), Parulidae (New World warblers) (53) Cuculidae (Cuckoos) (50), and Thraupidae
(Tanagers) (40).
Conservation Status
Lists of globally threatened and near-threatened African-Eurasian migratory landbird species,
species with decreasing global population trends, and species with increasing, stable or
unknown global population trends are available in Annex 3 of AEMLAP. See Table 14 for the
conservation status of landbird species according to the IUCN Red List.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
85
Table 14: Landbird conservation statuses according to IUCN
Threatened
CR
EN
VU
Subtotal
threatened
NT
LC
DD
Accipitriformes
13
20
24
57
31
162
1
Caprimulgiformes
12
19
23
54
39
498
10
Charadriiformes
11
15
25
51
45
281
2
Columbiformes
13
18
34
65
50
237
1
Coraciiformes
4
0
13
17
26
142
1
Cuculiformes
2
2
8
12
8
131
0
Falconiformes
0
2
6
8
6
50
0
Galliformes
11
20
45
76
48
183
0
Gruiformes
9
11
29
49
14
104
2
Passeriformes
91
196
348
635
515
5450
24
Piciformes
3
9
14
26
43
414
1
Strigiformes
4
11
28
43
27
167
2
Key: CR (critically endangered), EN (endangered), VU (vulnerable), NT (near threatened), LC
(least concern), DD (data deficient)
Figure 30 Sociable Lapwing (Vanellus gregarious). Photo: Sergey Dereliev.
Distribution and Habitat
A common pattern is for migratory landbirds to breed in the temperate, boreal, or Arctic biomes
of the northern hemisphere during the boreal summer, and then to spend the non-breeding
season in the warmer biomes of the northern hemisphere temperate and subtropics and
northern and southern hemisphere tropics, with fewer species migrating very long distances
to reach the temperate zones of the southern hemisphere during the austral summer (Kirby et
al., 2008). Intra-
convergence oscillates annually from the Tropic of Cancer to the Tropic of Capricorn and back
again. In the southern hemisphere, the predominant migratory pattern in the southern
hemisphere is for birds to breed in the temperate latitudes of South America, Africa and
Australasia, and to migrate to the tropics and subtropics in the austral winter (Kirby et al.,
2008).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
86
For a detailed review of bird flyways see UNEP/CMS (2014), noting that a flyway is defined
or a distinct population of a given migratory species completes all components of its annual
cycle (breeding, moulting, staging, non- breeding etc.). For some species and groups of
S, 2014). It should be noted that even
though flyways group birds into generalised strategies and patterns of movements, broad front
movements may characterise these strategies and patterns. Furthermore, flyways do not
necessarily capture all patterns and strategies, with some populations and species traversing
multiple flyways.
The East Asian-Australasian Flyway (EAF) is bounded by the 90
th
meridian on the west and
the Pacific Ocean on the east (Yong et al., 2021). It includes boreal, temperate and tropical
flyways. It also has the most threatened species. Two main migratory corridors are recognised
tern Russia and Japan to the
and continental Southeast Asia.
Over two billion birds travel the Afro-Palaearctic bird migration system annually, comprising
over 100 species, over 80% of which are songbirds and near-passerine birds (Briedis et al.,
2019; Moreau, 1972). Long-distance migrants travel between European breeding and sub-
Saharan non-breeding grounds via two broadly defined flyways (the Western flyway and the
Eastern flyway) which converge between 10 and 20
o
E in Central Europe (Briedis et al., 2019).
Billions of landbirds migrate in North America annually. In the spring, 2.5 billion migratory
landbirds migrate in and out of the contiguous USA at the south of the country and 2.7 billion
at the north (Dokter et al., 2018). Three flyways have been identified in North America: a
western flyway located to the west of 103
o
W longitude and an eastern flyway and a central
flyway which are interrelated and located east of 103
o
W (La Sorte et al., 2014). Most New
World landbirds spend the winter in tropical or south temperate latitudes with the majority
staying north of the equator in Mexico, the West Indies or northern Central America, but with
some species travelling further into southern South America (Faaborg et al., 2010). As north
temperate breeders move northwards during the Nearctic spring, birds from the temperate
zone of South America move northward to avoid the austral winter. An average of 2.1 billion
birds migrate through the Gulf of Mexico in the spring to reach their Nearctic breeding grounds
(Horton et al., 2019b). Smaller spatial migrations also take place, for example with birds that
breed at high elevations migrating to lower elevations before winter (Faaborg et al., 2010).
Some lowland tropical species also migrate according to annual wet and dry cycles.
Migratory landbirds need suitable habitat for feeding, resting or moulting during their migration
(Newton, 2008). Different strategies are used to move between habitats while migrating. Some
birds require closely interspersed habitats, others fly greater distances to pass ecological
barriers such as over expanses of sea, desert or mountains, before reaching the next relevant
habitat, whilst others undertake long-distance flights from one hemisphere to another.
Appropriate feeding areas before departure and upon arrival as well as appropriate stop-over
sites are essential for migrating birds.
Effects of Artificial Light on Migratory Landbirds
It has long been known that light at night has powerful effects on migratory birds. For example,
century-old records exist of extensive lighthouse collisions, and hunting, tourism and research
have systematically employed light to capture birds (Harvie-Brown, 1880; Beadnell, 1937;
Jones and Francis, 2003). For example, perhaps the most well-known capture site of landbirds

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
87
is Ngulia Lodge in Kenya where floodlights were used to illuminate wildlife for tourism from the
1960s, before a programme of mist-netting and banding began which has banded nearly a
million migrating birds (Moreau, 1972; Watson, 2017).
Seabirds and migratory shorebirds are recognised as needing protection from light pollution
and they are covered by Appendices G and H of these Guidelines. Migratory landbirds are
also at risk from the negative impacts of artificial light at night with additional threats such as
collisions with buildings and, therefore, this appendix has been developed to provide further
advice.
Of the 298 migratory landbird species considered by Cabrera-Cruz et al. (2018), all but one
had light pollution in their geographic distribution range. Light pollution was relatively greater
within the passage ranges of nocturnally migrating landbirds compared to their distribution
ranges during the other phases of their annual cycle. Long distance migrants often leave from
and arrive in areas with low levels of light pollution, but during migration they frequently cross
areas with high urban development and light pollution. Horton et al. (2019a) found that in the
eastern USA autumn migration routes take landbirds over areas with more light pollution than
spring routes, whereas on the west coast of the USA, landbirds have higher exposure during
spring migration. Chicago, Houston and Dallas are the US cities where landbirds were most
exposed to anthropogenic light, regardless of season.
Flight routes of landbirds can be affected by ALAN either through attraction or, conversely,
, evident in numerous publications
including, recently, Van Doren et al (2017). The illumination of buildings with indoor and
outdoor lighting, as well as contributions from other structures such as artistic installations,
arenas, stadia, towers and billboards, can create a skyglow visible for tens to hundreds of
kilometres. Numerous birds (e.g. Bruderer et al., 2018; dashboard.birdcast.info) fly between
ground level and 700 metres above ground level, bringing them into close proximity to
attractive and disorienting light, and so, too, to structures with which they can collide (e.g. Van
Doren et al., 2021; Korner et al., 2022; Lao et al., 2023). Although previous research
highlighted age, migration, phenology, and often specific weather conditions associated with
collisions, particularly those associated with poor visibility and increased moisture in the air
(Elmore et al., 2021a; Riding et al., 2021; Colling et al., 2022; Lao et al., 2023; Scott et al.,
2023), clear air conditions are also associated with large attraction events when illumination
can expand for many tens of kilometres (e.g. Van Doren et al., 2017). Birds aggregate in large
numbers, circle and or decrease their flight speeds, remain in close proximity to light, which
increases the risks of collision and predation, and alter social behaviours (e.g. flight calling)
Van Doren et al., 2017; Winger et al., 2019).
Light attracts and disorients nocturnally migrating birds. Numerous studies highlight these
behavioural responses, including attraction and disorientation (e.g., aggregation, circling) and
disproportionate occurrences of birds in urban areas because of these behaviours, as well as
enormous numbers of dead birds due to collisions (Allen, 1880; Gastman, 1886; Cochran and
Graber, 1958; Evans Ogden, 1996; Longcore and Rich, 2004; Gauthreaux and Belser, 2006;
Spoelstra and Visser, 2013; La Sorte et al., 2017; McLaren et al., 2018; Winger et al., 2019;
La Sorte and Horton, 2021; Korner et al., 2022).
Mechanisms by which light affects landbirds
The mechanism which causes birds to aggregate in light is not fully understood and could be
due to magnetoreception disruption, misinterpretation of natural light cues, or due to an effect
Many light orientations affect birds. Upward pointing lighting and lights on tall buildings or
structures affect flight behaviour of night migrating landbirds (Cabrera-Cruz et al., 2021). Van

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
88
Doren et al. (2017) found that birds reacted to vertically-oriented light beams up to 4km above
the ground, importantly during clear ).
However, low-rise lights which point downwards can also have an impact on landbird
behaviour, causing them to turn horizontally or vertically within their flight paths (Cabrera-Cruz
et al., 2021).
Collision is a major concern when considering how ALAN affects migrating landbirds. A study
in Minneapolis, Minnesota found that lighting area and lighting proportion had a statistically
significant positive association with the number of landbird collisions at building façades (Lao
at night were important predictors of collisions, and that lighting area in particular was a better
predictor than glass area, glass percentage, and the maximum and average sizes of glass
influences bird collision fatalities in spring, as well as the number of species colliding overall
and i
attracted birds (mainly passerines) which subsequently collided with the building (Korner et
al., 2022). When the façade illumination was reduced, there was a significant reduction of
casualties. Unlike in many other studies, Korner et al. searched for casualties throughout the
night. They found that the majority of casualties happened at night, and not, as sometimes
assumed, in the early morning.
In the USA, between 365 and 988 million birds die each year due to collisions with buildings
and other human-built structures (Loss et al., 2014). Most of these deaths involve collisions
with buildings, particularly windows, and involve migrating native species (Elmore et al.,
2021b). The number of fatal bird collisions in the USA, Canada and Mexico is greatest for
migratory, insectivorous and woodland-inhabiting species (Elmore et al., 2021a). Of the birds
killed at communication towers in the USA and Canada, the majority are neotropical migrants
and 97.4% of birds killed are passerines, mostly warblers (Parulidae, 58.4%) (Longcore et al.,
2013). The most visible publications regarding building collisions relate to sampling in the
eastern USA during migration, and this bias is represented in the species which have been
identified as being particularly vulnerable to collisions (Loss et al., 2014). The timing of
more likely to suffer a collision than diurnal migrants (Nichols et al., 2018; Colling et al., 2022).
Within species, juveniles are over-represented, possibly due to lack of experience (Colling et
al., 2022).
Collisions may be more likely to take place at night in some areas during certain weather
conditions, for example when there is low cloud, fog or rain and birds are flying at lower
altitudes (Newton, 2007; Elmore et al., 2021b). Studies at offshore installations have found
that migrating passerines are more attracted to artificial light on overcast nights (Poot et al.,
2008; Rebke et al., 2019). The attraction effect of blue light at narrow passes in mountain
ranges in Southwest China was also greatest during nights with fog and headwinds with the
majority of the birds captured being passerines (456 of 705 birds) with herons, cuckoos, doves
and crakes also caught (Zhao et al., 2020). However, attraction even in clear sky conditions
(e.g. Van Doren et al., 2017) and imperfect mortality sampling suggests more information is
needed to clearly define under what conditions such collisions are more prevalent.
Some studies suggest that glass or window area may be more of an influencing factor than
lighted area. Based on casualties in the morning, Parkins et al. (2015) concluded that the
amount of glass on a building façade next to an urban park in New York may have a greater
effect on collisions than the amount of light emitted from the façade. Configuration of glass on
building façades may also be relevant during daytime, with reflections of nearby habitat
confusing birds (Schneider et al., 2018). Potential solutions include physical barriers which
cover windowpanes, the use of patterns in manufactured panes which are visible by birds
when viewed from outside or the application of adhesives which uniformly cover the glass

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
89
surface (Klem, 2008). Adhesives or decals need to be applied so that the gaps between them
are small (5-10cm / 2-4 inches). There is also a potential for the use of ultraviolet (UV)
coverings which are visible to birds but not humans. The use of single items such as a falcon
silhouette has not been found to be effective. Further information is available here and here.
A study which looked at over 70,000 nocturnal bird-building collisions in Cleveland, Ohio, and
Chicago, Illinois, in the USA found an interaction between flight calling and collisions where
landbirds had been attracted by ALAN (Winger et al., 2019). This may be because calls from
individuals which have been attracted to the light cause more birds to be attracted to the lit
area. Flight-calling behaviour is, therefore, an important predictor of collision risk (Winger et
al., 2019). Gillings and Scott (2021) found that nocturnally migrating thrush call rates in the
UK were higher over bright urban areas compared to darker villages. The mechanisms
involved are not clear whether birds are altering their routes to pass over lit areas, whether
they fly at lower altitudes over lit areas, increase their call rate over lit areas or remain longer
over lit areas (Watson et al., 2016). The effects of artificial light need to be considered when
comparing abundance across sites (Gillings and Scott, 2021).
Positive phototaxis is not the only reaction observed in migrating landbirds. Sometimes
migrating birds may avoid brightly lit areas (negative phototaxis). Experimental evidence
shows that bright beams lead to aversive shifts in direction, speed and altitude of migratory
birds (Bruderer et al., 1999). Some observational data support these findings. For example,
birds stopping over in Sabancuy and Cancun in the Yucatan peninsula, Mexico during their
migration avoided bright lights in spring during stopover (Cabrera-Cruz et al., 2020). In
Cancun, more birds stopped over in areas away from bright lights in the fall/autumn too though
there were still relatively high bird densities closest to bright areas. Cabrera-Cruz et al. (2020)
proposed that naïve and ALAN-attracted birds are selected out during their southward
migration in the fall and that a higher proportion of ALAN-resistant individuals return north in
the spring.
When birds are attracted to or repelled by ALAN during their migration, this could result in
migration being less efficient, and time and energy requirements to complete it are increased
(La Sorte et al., 2017; Rebke et al., 2019). If birds are attracted to urban areas, they may find
less suitable habitat for foraging as well as increased hazards such as predators (cats, dogs,
rats etc.) and collision risks (La Sorte et al., 2017).
Effects of ALAN on timing of migration and other seasonal behaviours are expected to be
substantial especially through disruption of biological clocks. For example, songbirds are
known to misinterpret ALAN as a longer photoperiod (Dominoni and Partecke, 2015),
associated with continental-scale advancements of laying dates across the USA (Senzaki et
al., 2020). Fewer studies have looked into effects of ALAN on migration timing. As predicted
from interpreting ALAN as a longer photoperiod, purple martins (Progne subis) that
experienced the highest number of nights with ALAN at their overwintering sites were found
to depart for their spring migration an average of 8 days earlier than those that experienced
no artificial light (Smith et al., 2021). They also arrived 8 days earlier at their breeding sites. It
is possible that night migrants that synchronise migration to the lunar cycle suffer similar
mistiming (Norevik et al., 2019). Delayed or early arrival at breeding or wintering grounds
caused by ALAN mean that survival and reproductive success could potentially be impacted
if there is a mistiming with environmental conditions.
Migratory and non-migratory birds also experience other effects of ALAN. Dependent on their
anatomy, they can sometimes benefit from artificially extended feeding opportunities, but birds
also incur further physical costs (Lebbin et al., 2007; Sanders et al., 2021; Senzaki et al.,
2020). These include impaired physiology and health because of disruption of the circadian
clock (e.g., Dominoni et al., 2013; Kernbach et al., 2020). Because long-distance migrants are
typically insectivores, they may also be particularly affected by massive declines in insects

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
90
which have been linked to ALAN (Owens et al., 2020). Attraction to ALAN could also negatively
impact nocturnally migrating landbirds by increasing their exposure to air pollution and fine
particulate matter (PM
2.5
) in particular (La Sorte et al., 2022). Of the three flyway systems
assessed (Americas, Africa-Europe and East Asia-Australasia) by La Sorte et al. (2022), the
East Asian-Australasian flyway had the strongest ALAN-PM
2.5
correlations within its regions of
passage.
Wavelength, intensity and direction
Landbirds are able to differentiate between and, potentially, react differently to different colours
(Rebke et al., 2019). Most landbirds have a visual spectrum which extends into the UV range,
as well as non-visual light perception, for example in the brain (Falcón et al., 2020). In addition
navigation. Magnetoreception presumably involves two pathways. One, presumably located
in the beak, uses magnetite as a compass substrate. The second one is light-dependent and
likely involves a protein, probably a cryptochrome, which is activated by blue light and is
located in the retina including in cones (Günther et al., 2018; Pinzon-Rodriguez et al., 2018).
In caged birds, monochromatic red light and darkness led to disorientation. However, free-
flying migrants successfully navigate at night, for example by the stars or possibly using the
light-independent magnetic compass.
Some studies have attempted to determine whether landbirds react to different light
wavelengths. In a study by Poot et al. (2008) nocturnally migrating birds were recorded as
being attracted to and disoriented by red and white light (with visible long-wavelength
radiation). Poot et al. (2008) found it hard to identify birds to a species level but identified that
they were mostly passerines including thrushes and smaller songbirds but also some
shorebirds, ducks and geese. Gauthreaux and Belser (2006) also reported that migrating birds
were attracted by longer wavelengths in the light emitted by ceilometers and that when longer
wavelengths were filtered out so that mainly UV light was emitted, attraction was greatly
reduced. They also reported greater disorientation caused by red lights than white strobe
lights.
Poot et al. (2008) reported that the birds in their study were less disoriented by blue and green
light (containing less or not visible long wavelength radiation). Evans (2010) questioned Poot
sample sizes and the lack of information about migration density. Evans (2010) recommended
further rese
safer because birds are theoretically much less sensitive to it visually at night and fewer birds
found that nocturnally migrating birds (mostly passerines) were rarely attracted to long-
wavelength red light. In their study, short-wavelength blue light caused the strongest
phototactic response. Rebke et al. (2019) found that significantly more passerines were
attracted to continuous green, blue and white light than to red light at an offshore installation.
Recently, Adams et al. (2021) reviewed research looking at the effects of ALAN on birds and
found that most studies had looked at Passeriformes followed by shorebirds and seabirds.
They highlighted the need for further research into how different coloured lights affect birds as
they found that most studies had focused on red light.
Flashing lighting (on aviation obstruction towers, for example) causes less aggregation of
nocturnally migrating songbirds than continuous lighting (Evans et al., 2007). Communication
towers lit only with flashing lights (white, flashing strobes; red, strobe-like lights or red, flashing,
incandescent lights) caused fewer bird mortalities when compared to those towers lit with a

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
91
combination of red, flashing lights and red, non-flashing lights (Gehring et al., 2009). Rebke et
al. (2019) found that when crossing the sea when stars were not visible, more nocturnally
migrating passerines were drawn to continuous lights than blinking lights.
Light intensity may be relevant as well as wavelength (Cohen et al., 2021), although
nocturnally migrating passerines flying over the sea are known to have been attracted by even
relatively low intensity sources of light (Rebke et al., 2019).
Mitigation
Since 1993, Fatal Light Awareness Program (FLAP) Canada has worked to reduce deadly
landbird collisions with buildings. In 1995, FLAP Canada launched the first
initiative with World Wildlife Fund Canada with building managers turning off their lights at
night to help migrating landbirds. This campaign has led to many other similar initiatives across
North America and a number of cities and organisations have produced guidelines about how
to reduce light pollution for landbirds and how to improve building design to prevent collisions
(see Associated Guidance below). In the USA, bird collision deterrence is included as a credit
al Design)
system which determines standards of sustainability for the commercial, residential and
institutional building industries.
Turning off exterior lights has dramatic and immediate positive effects in reducing behavioural
responses of birds to light and allowing birds to resume typical migratory behaviours (Van
Doren et al., 2017). Tribute in
Lightexceeded magnitudes 20 times greater
ehavioural disruptions
disappeared when lights were extinguished, highlighting that the removal of light during nights
with substantial bird migration is a viable strategy for minimising potentially fatal interactions
involving ALAN, structures, and birds.
For some species, nights of intense migration can be forecast so mitigation measures can be
focussed on times when there is a greater risk for migratory landbirds. Weather radar can be
used to predict migration and, therefore, mitigation can be targeted at particular periods of
time, and/or specific weather conditions (Elmore et al., 2021b). Horton et al. (2021) found that
the majority of total migratory passage (54.3%) took place on 10% of nights for each season
in the contiguous United States and, therefore, recommended that using near-term ecological
-
BirdCast
(see here, here, and here) and other specific migration alerts, could all be informed
by radar data and could take into account particular periods of the night depending on
migration speeds and weather conditions (Elmore et al., 2021b).
Environmental Impact Assessment of Artificial Light on
Migratory Landbirds
As a minimum, infrastructure with artificial lighting that is externally visible should have Best
Practice Lighting Design implemented. An EIA should be undertaken where there is important
habitat for landbirds within a relevant distance of a project. The following sections step through
the EIA process with specific consideration for landbirds.
Where artificial light is likely to affect migratory landbirds, consideration should be given to
mitigation measures at the earliest point in a project and used to inform the design phase.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
92
It is important to recognise the spatial and temporal characteristics of migratory corridors for
some migratory landbird species. Species typically use established migratory pathways at
predictable times and artificial light intersecting with an overhead migratory pathway should
be assessed in the same way as for ground-based populations.
Associated guidance
• AEWA Plan of Action for Africa 2019-2027
• East Asian – Australasian Flyway Partnership 2019-2028 Strategic Plan
• Bird Cast
• FLAP
•
• Bird-Friendly Building Design
• Approved conservation advice.
Qualified personnel
Lighting design/management and the EIA process should be undertaken by appropriately
qualified personnel. Lighting management plans should be developed and reviewed by
appropriately qualified lighting practitioners who should consult with appropriately trained
ornithologists and/or ecologists.
Step 1: Describe the project lighting
The type of information collated during this step should consider the effects of artificial light on
migratory landbirds. Landbirds are susceptible when active at night while migrating, foraging
etc. The location and light source (both direct and skyglow) in relation to breeding,
overwintering and stopover sites as well as migration route should be considered.
Step 2: Describe migratory landbird population and behaviour
The species, life stage and behaviour of landbirds in the area of interest should be described.
This should include the conservation status of the species; abundance of birds; how
widespread/localised the population is; the regional importance of the population; and
seasonality of landbirds utilising the area.
Where there are insufficient data available to understand the population importance of
demographics, or where it is necessary to document existing landbird behaviour, field surveys
and biological monitoring may be necessary.
Biological monitoring of landbirds
Any biological monitoring associated with a project should be developed, overseen and results
interpreted by an appropriately qualified biologist or ornithologist to ensure reliability of the
data.
The objectives of monitoring in an area likely to be affected by light include:
• Understanding the habitat use and behaviour of the population (e.g. migrating,
foraging, breeding). Important habitat for landbirds may need to be determined on a
country-by-country basis. Natura 2000 Sites in Europe and Key Biodiversity Areas
(KBAs) could be a starting point.
• Understanding the size and importance of the population

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
93
• Describing landbird behaviour prior to the introduction/upgrade of light
The data will be used to inform the EIA process and assess whether mitigation measures are
successful. Suggested minimum monitoring parameters (what is measured) and techniques
(how to measure them) are summarised in Table 15.
Table 15. Recommended minimum biological information necessary to assess the importance
of a migratory landbird population. Note: the information in this table is not prescriptive and
should be assessed on a case-by-case basis.
Survey Effort
Duration
References
Direct visual monitoring
during the day and at night
when observers can see
birds in illuminated
nocturnal scenarios.
Multiple times daily during
peak movement periods and
also throughout the year.
Van Doren and Horton, 2018
Loss et al., 2023
Bird Cast
Globam
Radar based analysis
using broad scale weather
surveillance radar
networks or smaller scale
monitoring.
This can happen every 5-10
minutes in terms of the
scans, and continuously for
at least 10% of nights during
migration season, ideally not
randomly assigned.
Acoustic monitoring to
detect nocturnally
migrating birds that are
vocal.
Same as above.
Thermal imagers to see
migration as it happens.
Similar to above.
Moon watching to
understand broad patterns
of movements.
Whenever available.
Analyses of individual
tracking devices
that provide detail on
nocturnal aerial
distributions of birds.
Whenever available.
Citizen science-based
mortality surveys.
Regularly during day and
night.
Additional migratory landbird monitoring
• Monitor the population (during different seasons) to establish a benchmark for
assessing abundance before, during and after construction, and during operations to
detect project-related change.
• Measure nocturnal light levels at foraging sites and nocturnal roost sites before and
after the construction period of a project.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
94
• Monitor nocturnal roost sites using acoustic recording devices and/or infrared cameras
to determine nocturnal roost site use following the introduction of artificial light.
• Install camera traps at key locations to monitor collisions.
• Conduct nightly assessments of target lighting/areas to identify and collect birds which
have collided with infrastructure. Daytime surveys should also be carried out so that
collisions with glass can be identified. Conduct observations post-dusk and pre-dawn
with night vision goggles to assess activity/interactions.
• Track movement using land-based radar to determine existing flightpaths.
As a minimum, qualitative descriptive data on visible light types, location and directivity
should also be collected at the same time as the biological data. Handheld camera images
can help to describe the light. Quantitative data on existing sky glow should be collected, if
possible, in a biologically meaningful way, recognising the technical difficulties in obtaining
these data. See Measuring Biologically Relevant Light for a review.
Step 3: Risk assessment
The objective is that light should be managed in a way that the normal behaviours of migratory
landbirds are not disrupted. They should be able to undertake critical behaviours, such as
foraging and reproduction. Nor should they be displaced from important habitat. These
objectives should be considered in the risk assessment process.
In considering the likely effect of light on migratory landbirds, the assessment should consider
the existing light environment, the proposed lighting design and mitigation/management, and
behaviour of landbirds at the location. Consideration should be given to how the landbirds
perceive light. This should include both wavelength and intensity information and perspective.
To discern how/whether landbirds are likely to see light, a site visit should be made at night
and viewed from areas used by the birds. Consideration should be given to how birds will see
light when in flight. This could potentially be done using technology such as drones.
Step 4: Lighting management plan
This should include all relevant project information (Step 1) and biological information (Step
2). Maps of important areas for migratory landbirds and/or potential conflict areas should be
integrated into the planning process. The lighting management plan should outline proposed
mitigation. For a range of migratory landbird specific mitigation measures please see the
Migratory Landbird Light Mitigation Toolbox below. The plan should also outline the type and
schedule for biological and light monitoring to ensure mitigation is meeting the objectives of
the plan and triggers for revisiting the risk assessment phase of the EIA. The plan should
outline contingency options if biological and light monitoring or compliance audits indicate that
mitigation is not meeting objectives.
Step 5: Biological and light monitoring and auditing
The success of the impact mitigation and light management should be confirmed through
monitoring and compliance auditing and the results used to facilitate an adaptive management
approach for continuous improvement.
Relevant biological monitoring is described in Step 2: Describe migratory landbird population
and behaviour. Concurrent light monitoring should be undertaken and interpreted in the
context of how migratory landbirds perceive light and within the limitations of monitoring
techniques described in Measuring Biologically Relevant Light. Auditing, as described in the
lighting management plan, should be undertaken.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
95
Step 6: Review
The EIA should incorporate a continuous improvement review process that allows for
upgraded mitigations, changes to procedures and renewal of the lighting management plan.
Migratory Landbird Light Mitigation Toolbox
Appropriate lighting design/lighting controls and mitigating the effect of light will be site/project
and species specific. Table 16 provides a toolbox of management options relevant to migratory
landbirds. These should be implemented in addition to the six Best Practice Light Design
principles. Not all mitigation options will be practicable for every project. Table 17 provides a
suggested list of light types appropriate for use near important migratory landbird habitat and
those to avoid. The precautionary principles should be applied reducing ALAN to protect
migratory landbirds whenever possible.
Table 16: Light management options for migratory landbirds
Management Action
Detail
Turn out lights for as much of the
night as possible.
Exterior lights and interior lights that spill light
outside should be turned off for as much of the
night as possible to prevent negative impacts on
migratory landbirds.
Keep exterior lighting to a minimum.
Try to stay below legally permitted light levels with
outdoor lighting, noting that the desired
functionality can often be achieved with lower light
levels. Good visibility for humans depends on
avoiding too high contrasts between max. and min.
visible luminance. If visible luminance is reduced,
e.g. by shielding or suitable optical design, overall
lower illuminance levels can achieve an even
better visibility then higher illuminance levels, if the
visible luminance is also higher.
Keep lighting on land and at sea
(e.g. fishing boats, offshore wind
farms, oil and gas platforms) at a
minimum.
At sea, migratory landbirds regularly crash land on
vessels, so deck lights, navigational spot and
floodlights should all be kept to a minimum.
Use motion sensors to turn lights on
only when needed.
LEDs have no warm up or cool down limitations so
can remain off until needed and provide instant
light when required.
Avoid high intensity light of any
colour.
Keeping light intensity as low as possible will
minimise impact on migrating landbirds during
flight and at stopover sites.
Adapt spectra. As recommended in
Best Practice Lighting Design. Use
lights without blue, violet and
ultraviolet wavelengths where
possible.
Limit blue light and eliminate UV light.
Different scientific studies have come to varying
conclusions regarding how different coloured light
impacts birds and it is, therefore, important to refer
to the latest peer-reviewed literature regarding this
issue so that any new developments can be taken
into consideration.
Use flashing/intermittent lights rather
than continuous light if obstruction
lighting is needed.
Flashing lights can be used at offshore wind farms
or oil/gas platforms, communication towers and
other structures to reduce attraction/collisions.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
96
Management Action
Detail
If continuous light is needed use red
light.
Red light appears to attract fewer birds.
Aim lights downwards.
Aim light onto only the surface area requiring
illumination. Use shielding to prevent light spill into
the atmosphere and outside the footprint of the
target area.
Prevent indoor lighting reaching
outdoor environment.
Use fixed window screens or window tinting on
fixed windows and skylights to contain light inside
buildings. Where possible use black-out blinds,
shutters, curtains, localised task lighting, glass with
gl
Implement actions when birds are
likely to be present. This includes
peak migration periods (flyway
locations).
Most migration occurs within fixed time periods,
and according to local conditions. Within these
time periods, bird migration usually peaks on a
subset of nights. Migration forecasts can be based
on designated systems, such as radar, on other
information such as weather forecast, or on long-
term data sets including citizen science. Migratory
seasons should therefore be taken into
consideration when temporary lighting is being
planned e.g. at festivals.
Use curfews to manage lighting
during migration seasons e.g. lights
out from sunset to sunrise.
Extinguish as many exterior lights as possible and
block light spill from internal light sources during
curfews.
Do not use spotlights, searchlights,
floodlights and roof-top lighting.
Upward facing lights can affect migratory bird flight
behaviour and should not be used.
Turn off façade lighting during
migration seasons (especially
upward directed spotlights,
floodlights and roof-top lighting).
Upward facing lights can affect migratory bird flight
behaviour and should not be used at all but
particularly not when birds are migrating. If upward
facing lights are used for some reason, they
should be switched off when birds congregate in
them so that birds can disperse and continue their
migration. Downward facing façade lighting can
contribute to overall skyglow and light pollution and
should therefore also be avoided.
Define major flyways and stopover
sites as ALAN-free areas with the
goal to retain or restore night sky
luminance and the ambient lighting
levels to natural levels.
Such information can be based on long-term data
sets including citizen science, and on rapidly
increasing tracking information. Tracking
information is partly freely accessible in
repositories especially Movebank.org
Encourage building owners and
occupants to turn out all lights visible
outside during migration seasons
.
For more information see:
https://birdcast.info/science-to-action/lights-out/
https://www.audubon.org/lights-out-program
Publicise positive outcomes to encourage further
.
Monitor the
reductions in energy-usage, cost,
light emissions, bird collisions and
bird mortality.
Citizen scientists can be engaged, e.g. for
monitoring of casualties, and asked to provide
logistics information e.g. on costs (see Loss et al.,
2023).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
97
Management Action
Detail
Take into account bird migration
forecasts, where available, in the
management of artificial light at night
on flyways.
https://birdcast.info forecasts migration in the USA
and https://globam.science forecasts migration in
Europe and North America.
Develop migration forecasts globally
Key areas should be identified where migration
forecasts could help plan light pollution mitigation
actions (see Van Doren and Horton, 2018).
Take into consideration differences
in spring and fall/autumn migrations.
In some locations birds may be more exposed to
light pollution during one of their migrations.
Implement weather specific
measures.
In locations where more bird-building collisions are
recorded during foggy/overcast days, alerts should
be issued requesting lights to be turned off when
bad weather is forecast.
Integrate maps of hazard areas for
migrating landbirds into the planning
process.
Hazard areas are areas where large numbers of
birds are likely to come into contact with artificial
light at night.
Buildings with high levels of bird
mortality should apply appropriate
mitigation measures based on
expert advice building specific
mitigation methods.
Mitigation measures may vary from structure to
structure.
Minimise the reflection of vegetation
and water features on building
façades.
Birds perceive reflections of vegetation,
landscapes or sky to be real. By minimising
reflection on building façades, the risk of collision
can be lowered for birds which have been
attracted by light pollution to areas where bird-
building collision is a risk. Refer to existing building
guidelines for how to reduce collision risks for
Best Practices Glass.
Conduct regular surveys to monitor
landbird collisions during peak
migration periods.
programmes are being successful and to highlight
which buildings/locations need mitigation
measures.
Instruct monitors in methods of
caring for injured birds before they
can be transported to a wildlife
rehabilitator.
For example: https://flap.org/finding-an-injured-
bird/
Use other materials to remove need
for lighting.
Glow in the dark paths, reflective paints and
tapes and/or self-luminous materials for signs,
curbs, paths and steps can all be used instead of
installing lighting.
Table 17 Where all other mitigation options have been exhausted and there is a human safety
need for artificial light, this table provides commercial luminaires recommended for use near
migratory landbird habitat and those to avoid.
Light type
Suitability for use near migratory
landbird habitat
Low Pressure Sodium Vapour
High Pressure Sodium Vapour
Filtered* LED

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
98
Filtered* metal halide
Filtered* white LED
LED with appropriate spectral properties for
species present
White LED
Metal halide
White fluorescent
Halogen
Mercury vapour
only if a filter is applied to remove the problematic
wavelength light.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
99
Appendix J Bats
As they are nocturnal, bats are particularly susceptible to the impacts of light
pollution and a number of their behaviours including foraging, commuting, drinking,
roosting and migrating can be disrupted. Natural darkness should be maintained
wherever possible in areas where bats are present. Due to the great diversity
amongst species, if artificial light is present or is to be introduced, mitigation
measures should be site and species specific.
As many bat species are insectivores, consideration needs to be given to how their
insect prey is impacted by artificial light and how this, in turn, affects bats.
Bats are a highly diverse group of flying mammals within the order Chiroptera, with over 1,400
species, divided into 21 families. In recent decades many new species have been described;
over 270 new species have been described since 2005 (Frick et al., 2020). See Table 18.
Table 18: Families of Chiroptera (Burgin et al., 2020 and Simmonds and Cirranello, 2023)
Family
Number of species
Cistugidae
Wing-gland bats
2
Craseonycteridae
Hog-nosed bat
1
Emballonuridae
Sac-winged bats
55
Furipteridae
Smoky bats
2
Hipposideridae
Old World leaf-nosed bats
90
Megadermatidae
False vampire bats
6
Miniopteridae
Long-fingered bats
37
Molossidae
Free-tailed bats
132
Mormoopidae
Ghost-faced bats
18
Mystacinidae
Short-tailed bats
2
Myzopodidae
Sucker-footed bats
2
Natalidae
Funnel-eared bats
11
Noctilionidae
Bulldog bats
2
Nycteridae
Slit-faced bats
15
Phyllostomidae
New World leaf-nosed bats
227
Pteropodidae
Old World fruit bats
199
Rhinolophidae
Horseshoe bats
110
Rhinonycteridae
Trident bats
9
Rhinopomatidae
Mouse-tailed bats
6
Thyropteridae
Disk-winged bats
5
Vespertilionidae
Evening bats
523
Chiroptera is the second most speciose mammalian group and yet there remain key
challenges in understanding its taxonomy, which to some extent remains in flux, and the
ecological roles played by bats (Kruskop, 2021). Bats exhibit a wide variety of lifestyles for
example in their foods, with many eating insects and others eating fruit and nectar and their
wide range of behaviours and habitats makes it challenging to provide guidelines that reduce
the effects of artificial light at night for all species. Hence, the overarching recommendation
that specific guidelines should be developed on a local basis to suit the species and habitat
concerned.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
100
Figure 31 Greater horseshoe bat (Rhinolophus ferrumequinum). Photo: Paulo Barros.
Conservation Status
According to the IUCN Red List 219 species of Chiroptera are considered threatened (either
Critically Endangered, Endangered or Vulnerable). Ninety-four species are Near Threatened,
773 are Least Concern and 237 are Data Deficient. Many of the species which have only been
described in recent years have still not been classified by the IUCN (Frick et al., 2020).
Bats are protected under various international treaties and agreements including the Habitat
Directive of the European Union, the Convention on the Conservation of Migratory Species of
Wild Animals (CMS, Bonn 1979) and the Agreement on the Conservation of Populations of
European Bats (EUROBATS) which was set up under CMS in 1994. At the 8
th
session of the
Meeting of the Parties of EUROBATS in 2018, Resolution 8.6 Bats and Light Pollution was
adopted.
The following Chiroptera species are listed on Appendix I of the Convention on International
Trade in Endangered Species of Flora and Fauna (CITES, Washington 1973): Acerodon
jubatus, Pteropus insularis, Pteropus loochoensis, Pteropus mariannus, Pteropus molossinus,
Pteropus pelewensis, Pteropus pilosus, Pteropus samoensis, Pteropus tonganus, Pteropus
ualanus, Pteropus yapensis. Acerodon spp. (except the species included in Appendix I) and
Pteropus spp. (except the species included in Appendix I and Pteropus brunneus which is not
included in the Appendices) are on Appendix II. Platyrrhinus lineatus (Uruguay) is on Appendix
III.
In 2015, a Letter of Intent Related to Efforts to Promote Conservation of Bats in the United
Mexican States, the United States of America and Canada was signed.
Distribution and Habitat
Bats can be found on all continents except Antarctica, and they are particularly abundant in
the tropics and some temperate ecosystems (Voigt and Kingston, 2016). The highest
concentrations of species are in tropical America, tropical Africa and Southeast Asia (the
Indochina, Sumatra, Borneo region) (, 2005). Southeast Asia is a global hotspot with

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
101
at least 388 species (Yoh et al., 2022). Europe and North America have fewer than 50 species
each (Ulrich et al., 2007).
Important habitat for bats
Bats require suitable habitat for roosting, commuting, foraging, drinking and other key
behaviours. Habitat choice is species-specific and some bats will travel further than others to
find their preferred habitat (Bat Conservation Trust, 2023a). Insectivorous bat species, for
example, may have a preference for foraging at waterways, in woodlands or over grasslands
and farmlands. Foraging areas around and near maternity roosts are important habitat areas
for bats particularly because the energy demands for pregnant and lactating females are high
(Kyheröinen et al., 2019). The commuting areas linking foraging areas and maternity roosts
are also important. Commuting routes may follow natural landscape features such as rivers,
hedgerows and tree-lined footpaths (Bat Conservation Trust, 2023b).
Regarding habitat for pteropodid species, many are reliant on plantations and gardens
whereas only 11% are dependent on primary vegetation only, and fifteen species use urban
landscapes to forage (Aziz et al., 2021). This means that the majority are using habitats which
could potentially bring them into conflict with humans.
Effects of Artificial Light on Bats
As largely nocturnal mammals, bats are particularly susceptible to disruption from ALAN.
Roosting, emerging, commuting, foraging, drinking, swarming, migrating and mating
behaviours could all potentially be disrupted (see Glossary for definitions of roost, commuting
route and swarming). More examples are provided below, and bats are considered under two
broad headings dividing them into principally insect feeding and fruit/nectar feeding species
(including pteropodid and phyllostomid bats).
Trawling bats which feed on insects and small fish are also known to be impacted by artificial
light at night (Haddock, 2018). Bats which feed on blood (subfamily Desmodontinae) are not
considered here due to a lack of information regarding how they are impacted by ALAN. Future
research should look at these groups.
This is not meant to be an exhaustive review but is intended to highlight what is known of
some of the concerns and hence the rationale for addressing light pollution for bat species. It
should be noted that most of the available information on the effects of artificial light on bats
comes from temperate areas, where the vast majority of the species are in the family
Vespertilionidae, all insectivores, and very little or no work has been done in tropical areas.
More work is being done, but it will be years before we have a clear view of the general
patterns. Thus, a precautionary approach should be adopted until we know more.
Effects of Artificial Light on Insectivorous Bats
Many bats rely on invertebrates for their food. A large part of understanding behaviours of
insectivorous bats around artificial lights requires understanding how their insect prey is
attracted to lights (Voigt et al., 2018a).
Mechanisms by which light affects insects
Eisenbeis (2006) reviewed the different ways in which insect behaviour is affected by artificial
lights including

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
102
light and die immediately, it may orbit the light until caught by a predator or until it dies from
exhaustion, or it may manage to move away from the light for a while but as it remains inactive
because of exhaustion or because it is dazzled by the light it is, therefore, at greater risk of
occurs when streetlights prevent insects from following
their original foraging or migratory route, subsequently causing them to get trapped by the
or migrating, leading to their deaths and, potentially, causing a reduction in the local
population. As well as attraction, lights can have other impacts on nocturnal insects, such as
their visual systems being desensitised, a loss of ability to recognise objects in their
environment and temporal or spatial disorientation (Owens and Lewis, 2018). For insects
such as mayflies that normally oviposit on water cued by how light is polarised by the water
surface to lay their eggs, artificial light can lead to them being drawn to asphalt surfaces and
ovipositing on roads and bridges which are artificially lit (Szaz et al., 2015).
The strength of attraction also depends on the type of lamp used and the wavelengths it emits.
Spectral composition may be more important than light intensity for insects (Longcore et al.,
2015) with UV emitting lights attracting more insects (Barghini and Souza de Medeiros,
2012). However, Bolliger et al. (2020) found that intensity could also be relevant and that the
more light emitted by LED streetlights in Switzerland, the more insects were caught in insect
traps. Heteroptera were particularly sensitive to light levels and the dimming of lights seemed
to benefit them. Caution is needed when using how many insects are attracted to a light to
activity and, therefore, attract fewer insects (Boyes et al., 2021). The distance from which
insects can be attracted to lights varies depending on background illumination and the height
of the artificial light (Eisenbeis, 2006). During the full moon, for example, fewer insects are
attracted to artificial lights.
There may be differences between insect orders in terms of what kind of light they are attracted
to (Desouhant et al., 2019). More Coleoptera were attracted to a high-pressure sodium (HPS)
light than an LED, whilst Diptera were more diverse around LEDs (Wakefield et al., 2018).
Different families of Lepidoptera respond differently to light. For example, shorter wavelength
lighting attracted more Noctuidae than longer wavelength lighting (Somers-Yeates et al.,
2013). Geometridae were attracted by both wavelengths. Certain moth species or families
might be more attracted by UV light than others, with those attracted to UV-emitting lamps
dying from either exhaustion or predation, while others are less affected (Straka et al., 2021).
There is concern that artificial light at night, alongside other drivers including habitat loss,
pesticide use, invasive species and climate change, is contributing to the rapid decline of
insects worldwide (Owens et al., 2020). This decline in insects has many implications
including, of course, for insect predators such as bats (Voigt et al., 2018a).
Some actions which are recommended for reducing obtrusive light, light spill and skyglow,
such as shielding of lights, are not sufficient to prevent insects in the immediate area of a light
from being affected (Owens et al., 2020). Insect conservation requires the limiting of lighting
to necessary areas, using the lowest safe intensity and reducing the number of fixtures
installed especially close to ecologically vulnerable areas. Seasonal approaches may also be
appropriate in some cases. How insects are affected by polarization and flicker rate needs
further investigation.
Impacts of artificial light on bat foraging activity
Presence of insects under lights may attract some species of foraging bats, particularly fast-
flying aerial hawking species which forage in open areas (e.g. genera Eptesicus, Nyctalus and
Pipistrellus) (Stone et al., 2015; Lacoeuilhe et al., 2014). Eptesicus species in Sweden have

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
103
been found to benefit from the increase in prey available at bright streetlights (Rydell, 1992).
However, a short-term increase in availability of insects at light sources may cause insect
populations to decline in the long term and thereby reduce food availability for bats. Bats
which forage near artificial lights may put themselves at greater risk of predation.
More light-averse species such as Myotis, Plecotus or Rhinolophus may avoid foraging near
both bright and dimmed streetlights and could, therefore, lose foraging sites when artificial
light is installed (Stone et al., 2015; Luo et al., 2021). In Missouri, USA, Eastern red bats
(Lasiurus borealis) actively forage around lights, particularly just after sunset, whereas other
species, including big brown bats (Eptesicus fuscus) and gray bats (Myotis grisescens), avoid
lit areas (Cravens and Boyles, 2019).
These differences in foraging around artificial lights have led to bat species being divided into
light-sensitive or light-tolerant/light-exploiting species. However, Voigt et al. (2018a; 2021)
warned against such labels, as the reaction of a species to light can vary depending on several
factors according to the specific situation. They categorised the likely responses of the
different European bat genera in different situations as either an averse response, a neutral
response or an opportunistic response (see Voigt et al., 2018a). A recent review found that
how bats are impacted by ALAN depends on the context including the activity being
undertaken by the bat as well as on the specie
Although foraging guild can be a good generalisation, there can still be variation between
species and so care should be taken not to overly generalise. Precaution should still be taken
as light may not impact bat behaviour but may still impact physiology which can be difficult to
monitor or measure. All European species react sensitively to ALAN near their roosts and to
the illumination of drinking sites (Russo et al. 2017), possibly because of the increased risk of
predation. In areas where they commute or forage, effects are more varied.
ALAN can cause a shift in community composition and may disadvantage some species
(Russo et al., 2019; Seewagen and Adams, 2021). Introducing LED lighting to foraging habitat,
led to a decrease in the presence and activity of little brown bats (Myotis lucifugus) and a
reduction in activity for big brown bats and silver-haired bats (Lasionycteris noctivagans) in
Connecticut, USA, while red bats and hoary bats (Lasiurus cinereus) were not affected by the
lights (Seewagen and Adams, 2021). A study in Italy found that ALAN influenced niche
separation between common pipistrelles (Pipistrellus pipistrellus)
(Pipistrellus kuhlii), which are both streetlamp foragers (Salinas-Ramos et al., 2021).
pipistrelles used artificially lit areas more frequently than common pipistrelles. Species
richness in Peru decreased with artificial light intensity although eight species were recorded
using urban areas with high levels of ALAN (Mena et al., 2021).
For some species, ALAN along forest edges increases the probability of bats flying inside the
forest (Barré et al., 2021). This suggests that bats use landscape structures when they react
to light, for example to avoid predation. A study in Sydney, Australia found that bat activity
was higher in forest interiors compared to forest edges and that slower-flying species, which
are adapted to cluttered environments or with high characteristic echolocation call frequency,
were negatively affected by ALAN at the forest edge (Haddock et al., 2019a). The activity of
this group (which included Nyctophilus spp., Rhinolophus megaphyllus, Vespadelus vulturnus,
Chalinolobus morio and Miniopterus australis) decreased after high UV mercury vapour lights
were changed to low UV LEDs (Haddock et al., 2019b). The change to white LED streetlights
could therefore cause a decline in some insectivorous bat species abundance or changes in
community composition although this may depend on previous exposure to ALAN. Bats which
are relatively naïve to ALAN are more likely to show a reaction to it than bats in environments
with long-term sources of ALAN (Seewagen and Adams, 2021). In Singapore, for example,
where there are extremely high levels of light pollution (Falchi et al., 2016), changing HPS
streetlights for white LED streetlights did not influence bat activity (Lee et al., 2021). Species

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
104
that are less adapted to urban areas or areas with significant levels of ALAN may demonstrate
behavioural changes.
Bat activity was found to be impacted by a white LED lamp with a luminous flux of 6480 lm
(4000-4500 K) illuminating a cross section of river in the Central Italian Apennines (Russo et
Myotis daubentonii)
significantly increased under the light. Other species or species groups showed no significant
because Chironomidae and Ceratopogonidae numbers increased under the lit conditions,
mainly closer to the LED lamp, although the insect community over the water showed no
qualitative or quantitative changes. The bats, therefore, appeared to be avoiding the artificial
lighting.
Impacts of artificial light on bat roosts
Artificial lights near roost sites (locations used by bats for resting and socialising during the
day and, occasionally, during the night) can negatively impact bats by disrupting their
emergence activity and subsequently leading to reduced foraging opportunities because of a
reduction of time available for foraging as well as access to the peak availability of insects at
dusk (Stone et al., 2015; Voigt at al., 2018a). Rydell et al. (2017) found that bat colonies in
churches require one side or end of the church to remain unlit, preferably the part that is
nearest to surrounding tree canopies, so that bats can exit and return to the roost in safety.
Artificial light at a roost site can lead to increased predation particularly if bats are forced to
use an alternative, suboptimal exit (Stone et al., 2015). In some circumstances, light can force
a colony to abandon its roost. For example, a whole colony (1,000-1,200 females) of
Myotis emarginatus) abandoned a roost at a church in Hungary when
floodlights were installed (Boldogh et al., 2007). Colonies of brown long-eared bats (Plecotus
auritus) no longer roosted at several country churches in Sweden which had floodlights
installed (Rydell et al., 2017).
The presence of neutral white (broad spectrum of ~420-700 nm with peaks around 450 and
540-620 nm), red (spectrum between 620 and 640 nm with a peak around 630 nm) or amber
(spectrum between 580 and 610 nm with a peak around 597 nm) LED at a cave entrance
Miniopterus
schreibersii), long-fingered bats (Myotis capaccinii), Mediterranean horseshoe bats
(Rhinolophus euryale) and (R. mehelyi), with red LED having the
least negative effect (Straka et al., 2020). Rhinolophus species showed the strongest reaction.
Straka et al. (2020) investigated the short-term effects of light on cave-dwelling bats but
pointed out the potential for cumulative and long-term effects which could negatively impact
entire colonies.
Impacts of artificial light on commuting behaviour
When artificial light disrupts commuting routes, bats may have to use suboptimal routes
requiring increased flight time and energetic expenditure to arrive at their foraging grounds
(Stone et al., 2015). They may also be at greater risk of predation or exposure to wind and
rain. If no alternative route is available, then a colony may have to abandon its roost. Colony
losses of brown long-eared bats in Sweden may also be associated with artificial illumination
in their flight corridors (Rydell et al., 2021).
Vertical illuminance has been found to be a better predictor of bat activity than horizontal
illuminance, and so light orientation needs to be taken into consideration when assessing the
impacts of ALAN on bats (Azam et al., 2018).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
105
Some species are more likely to avoid light. For example, serotine bats (Eptesicus serotinus),
avoided lights at greater distances than other species (Azam et al., 2018). For these species,
light may be particularly likely to form a barrier and the placement of streetlights can impact
the movements of bats when they are foraging, for example. Bat activity in Sydney, Australia
has been shown to be higher in forest interiors compared to forest edges whether there is
artificial light at the forest edge or not (Haddock et al., 2019a). This highlights the importance
of maintaining connections or corridors between forest areas, especially forests in or close to
urban areas.
through culverts were not affected by artificial LED light of different colours (red, white, green)
with a light intensity of 5.0 ± 0.2 lx at the water level. The lack of response could have been
due to the experimental set-up, the low light levels used or the location of the culverts, which
passed under a road, and thereby the traffic noise may have deterred the bats more and
encouraged them to still use the culverts despite the addition of the LEDs. Bat reactions to
artificial light may be site specific and this highlights the importance of carrying out detailed
environmental impact assessments.
Impact of wavelength and light intensity on bats
Bats are impacted by lights of differing colours and intensities (Voigt et al., 2021) though
different species may be affected differently. During migration, soprano pipistrelles
(Pipistrellus pygmaeus) Pipistrellus nathusii) showed increased
activity when a red LED (with a dominant wavelength of 623 nm) was on, though this was not
associated with increased feeding, suggesting that the association of the bats with red light
was due to phototaxis (Voigt et al., 2018b). Spoelstra et al. (2017), however, found that
Pipistrellus, Plecotus and Myotis species were equally abundant in red illuminated areas
compared to a dark control, suggesting that there was no phototactic response when bats
were not migrating. Barré et al. (2021) found that Pipistrellus species were more likely to fly
inside a forest area when they were near red or white lights (compared to dark control areas)
and that the probability was greater for red light as the bats got closer to the light.
During migration, Pipistrellus did not show increased general activity at a warm-white LED
light source (dominant wavelength 581 nm), but they did demonstrate increased foraging
compared to the dark control (Voigt et al., 2018b). Spoelstra et al. (2017) found that Pipistrellus
species were more abundant around white and green lights while Myotis and Plecotus species
avoided them. Barré et al. (2021) also found that for Myotis and Plecotus, white lights had a
more significant effect than red lights, prompting them to fly inside a forest area when near the
lights. For Eptesicus and Nyctalus, bats were significantly more likely to fly inside a forest near
white light, though as they got closer to the lights, the probability of flying in the forest was
stronger for both red and white lights. Contrasting results in studies on light spectra could be
due to condition-dependent effects of ALAN on bats, for example before and during the
migration period when vision plays a more dominant role than echolocation (Voigt et al.,
2018b).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
106
Figure 32 Comparative light perception among different species groups
Note: Horizontal lines show a broad generalisation of the ability of humans and wildlife to perceive
different wavelengths. Dots represent reported peak sensitivities. Black dots for bats represent peak
sensitivities in an omnivorous bat, based on Winter et al. (2003); grey dots represent potential peak
sensitivities in bats, derived from Feller et al. (2009) and Simões et al. (2018). Figure adapted from
Campos (2017).
A study using dim, flickering UV lights (>400 nm) to deter bats from a wind turbine found that,
in fa
As there was not a significant increase in insect activity, it appeared to be the illuminated
surface of the wind turbine rather than the presence of insects which attracted the bats. Straka
et al. (2019) found that different species respond differently to the emission of UV
showed increasing activity with
an increasing number of UV emitting streetlamps whereas Soprano pipistrelles, and bats in a
group including the species Nyctalus and Eptesicus and the Particoloured bat (Vespertilio
murinus) (which could not be distinguished according to their echolocation calls) responded
negatively to mercury vapour and metal halide streetlights which emitted UV light.
Light intensity is important as well as spectrum. ALAN that is brighter than moonlight can
disrupt foraging and mating in bats as well as interfering with entrainment of the circadian
system (Voigt et al., 2018a). Increasing illuminance led to a decrease in bat activity and buzz
ratio with white LED lamps while the opposite effect was found with low-pressure sodium
(LPS) lamps (Kerbiriou et al., 2020). This could have been due to an associated greater
predation risk under stronger LED light which resembles daylight more than the light produced
by LPS. Different species are sensitive to different light intensities and some species avoid lit
environments, regardless of light intensity or spectrum (Kerbiriou et al., 2020). Illuminance
values lower than 1 lx had a negative effect on light-sensitive Myotis species, whereas
common pipistrelles and lesser noctules (Nyctalus leisleri), were most active between 1 lx and
5 lx. (Azam et al., 2018).
Even relatively short periods of artificial lighting can have a negative impact on bats so
reducing the period of time areas are lit is one mechanism to reduce impacts of ALAN, as well
as reducing light intensity. Boldogh et al. (2007) reported that for greater horseshoe bat
(Rhinolophus ferrumequinum), Geoffroy's Bat and lesser mouse-eared bat (Myotis
oxygnathus), even a one-hour lighting period after dusk can cause significant disruption in

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
107
the roost until it was totally dark. Azam et al. (2018) also found that the negative effect of ALAN
on Myotis species continued even after streetlights had been turned off.
Effects of Artificial Light on Fruit and Nectar Feeding Bats
Little is known about how tropical fruit and nectar feeding bats are affected by ALAN (Rowse
et al., 2016), although they tend to avoid areas which are well-illuminated (Hoyos-Díaz et al.,
2018). ALAN may prevent them from commuting and dispersing seeds leading to genetic
isolation of illuminated plants and other important impacts on ecosystems (Lewanzik and
Voigt, 2014). In areas where deforestation and light pollution are increasing, ecosystem
functioning may be seriously affected. Old World fruit bats are important pollinators and seed
dispersers with a number of species migrating seasonally to follow food resources (Aziz et al.,
2021). The straw-coloured fruit bat (Eidolon helvum), for example, plays an important role in
long-distance seed dispersal in sub-Saharan Africa (Aziz et al., 2021), one of the regions of
the world with the lowest levels of light pollution (Falchi et al., 2016). With increasing
urbanisation rates in some countries in Sub-Saharan Africa (UN, 2019), there is a potential for
increased light pollution to disrupt fruit bat activities with a knock-on effect for ecosystems.
Six times fewer great fruit-eating bats (Artibeus lituratus) and Jamaican fruit-eating bats (A.
jamaicensis) were captured in a secondary growth forest patch in Venezuela when HPS lamps
were installed (Hoyos-Díaz et al., 2018). Light pollution was also found to impact the intensity
with which great and Jamaican fruit-eating bats visited Ceiba pentandra trees in Yucatan,
Mexico (Dzul-Cauich and Munguía-Rosas, 2022). As pollinators, the reduction in bat
visitations could have impacted reproductive success for the trees but, in fact, this was not the
case and the artificial light (mean level 5.06 ± 0.86 lx with the highest level of 18.20 lx in this
study) had a direct and positive effect on C. pentandra reproductive success.
The time when Indian flying foxes (Pteropus giganteus) emerge from their tree roosts is highly
correlated with sunset and day length (Kumar et al., 2018). All individuals from a roost will
emerge within less than an hour, as will greater short-nosed fruit bats (Cynopterus sphinx)
rousette, Rousettus leschenaultii), emergence times are more spread out, with peak
emergence time varying according to the moon phase. Their flight activity is restricted to lower
light levels than tree-roosting species. Different species may, therefore, respond differently to
light pollution. Floodlights have been used successfully as a management tool to deter flying
foxes from roosting in particular trees in Queensland, Australia (State of Queensland, 2020).
In areas where flying foxes are protected under State legislation, therefore, it is necessary to
remove floodlights. Further investigation into how pteropodids respond to artificial light at night
is needed.
Green cover is important for plant-eating bats and so increasing the presence of vegetation
may be an important mitigation method to prevent any negative impacts from light pollution
(Dzul-Cauich and Munguía-Rosas, 2022).
Environmental Impact Assessment of Artificial Light on
Bats
As a minimum, infrastructure with artificial lighting that is externally visible should have Best
Practice Lighting Design implemented to reduce light pollution and minimise impacts on bats.
Where bat species are known to occur or are likely to occur in the area, an EIA should be
undertaken. The following sections step through the EIA process with specific consideration
for bats. In the European Union lighting within Natura 2000 sites should be subjected to a

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
108
specific assessment according to the Habitats and Species Directive (Council Directive
92/43/EEC).
Bats are susceptible to impacts from artificial light while roosting, commuting, foraging,
drinking, returning to roosts, swarming and migrating. The location of light sources (both direct
and skyglow) in relation to important habitats and features including roosts, caves,
hibernacula, swarming sites, associated flightpaths, commuting habitat, foraging areas and
water sources should be considered.
Associated guidance
• EUROBATS Guidelines for consideration of bats in lighting projects (Voigt et al.,
• Collins, J. (ed.) (2016) Bat Surveys for Professional Ecologists: Good Practice
Guidelines (3
rd
edition). The Bat Conservation Trust, London
• EUROBATS Guidelines for Surveillance and Monitoring of European Bats (Battersby
(comp.), 2017)
• The DarkCideS global database of bat caves and species contains information for
geographical location, ecological status and species traits (Tanalgo et al., 2021)
• The Action Plan for the Conservation of All Bat Species in the European Union 2018-
2024
Qualified personnel
Artificial lighting design/management and the EIA process should be undertaken by
appropriately qualified personnel. Lighting management plans should be developed and
reviewed by appropriately qualified lighting practitioners who should consult with appropriately
qualified biologists and/or ecologists.
Step 1: Describe the project lighting
The information collated during this step should consider the Effects of Artificial Light on Bats.
Describe the existing light environment and characterise the artificial light likely to be emitted
at the site. Information should include (but not be limited to): the location and size of the project
footprint; the number and type of artificial lights - their height, orientation and hours of
operation; site topography and proximity to bat habitat. This information should include
whether artificial lighting will be directly visible to bats or contribute to skyglow; the distance
over which this artificial light is likely to be perceptible; shielding or artificial light controls used
to minimise impacts; and spectral characteristics (wavelength) and intensity of artificial lights.
Step 2: Describe bat population and behaviour
Guidelines for
consideration of bats in lighting projects (Voigt et al., 2018a).
This should include a bat survey to find out which species occur in an area, where their roost
sites, feeding areas and commuting routes are. Good practice guidelines for bat surveys exist
and should be used. For example, Collins (2016) and Battersby (2017).
The species, behaviour and diet of bats roosting and foraging in the area of interest should be
described. This should include the conservation status of the species; population trends
(where known); how widespread/localised roosting for that population is; the abundance of

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
109
bats using the location; the regional importance of the population; the seasonality of roosting
and breeding; and foraging requirements and foraging range from roosting.
Where there are insufficient data to understand the population importance or demographics,
or where it is necessary to document existing bat behaviour, field surveys and biological
monitoring may be necessary. While bat colony roost sites may be known, commuting paths
are less likely to be known (Voigt et al., 2018a).
Biological monitoring of bats
Any monitoring associated with a project should be developed, overseen and results
interpreted by appropriately qualified and experienced personnel to ensure reliability of the
data.
The objectives of bat monitoring in an area likely to be affected by artificial light include:
• understanding the size and importance of the bat population(s);
• understanding any interspecies interactions (where multiple bat species are found at
the same site);
• identifying roosts, commuting routes and foraging and watering areas where artificial
lighting changes may occur; and
• describing bat behaviour at roost sites, foraging areas and commuting routes before
and after the introduction/upgrade of artificial light.
The data will be used to inform the EIA and assess whether mitigation measures have the
potential to be successful.
To understand existing bat behaviour it will be necessary to undertake monitoring (or similar
construction/lighting upgrades. Consideration should be given to monitoring a comparative
control/reference site to ensure observed changes in bat behaviour are related to changes in
the light environment and not broader climatic or other landscape-scale changes.
Artificial light can fragment and degrade bat habitat. Biological monitoring should include an
adequate population survey to determine if there are important bat populations.
A well-designed behavioural monitoring programme will capture the following both before and
after artificial lighting design implementation:
• Behaviour of bats at roost sites including location of roost used, type of roost used,
time of first emergence, time of return to roost, duration of rest and torpor.
• Foraging activity of bats including location and type of foraging sites, time spent
foraging, and prey availability.
• Commuting routes used by bats including location of commuting routes, time, and
duration of commuting behaviour.
Surveys should be designed in consultation with a quantitative ecologist/biostatistician to
ensure that the data collected provides for meaningful analysis and interpretation of findings.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
110
As a minimum, qualitative descriptive data on visible light types, location and directivity should
also be collected at the same time as the biological data. Quantitative data on existing skyglow
should be collected, if possible, in a biologically meaningful way, recognising the technical
difficulties in obtaining these data. See Measuring Biologically Relevant Light for a review.
Step 3: Risk assessment
The objective is that light should be managed in a way that the normal behaviours of bats are
not disrupted. They should be able to undertake critical behaviours, such as foraging,
commuting, and roosting. Nor should they be displaced from important habitat. These
objectives should be considered in the risk assessment process.
In considering the likely effect of light on bats, the assessment should consider the existing
light environment, the proposed lighting design and mitigation/management, and behaviour of
bats at the location. Consideration should be given to how bats perceive light. This should
include both wavelength and intensity information and perspective. To discern how/whether
bats are likely to see artificial light, a site visit should be made at night and the area viewed
from any known roosts and other key habitat. Similarly, consideration should be given to how
bats will see light when in flight. This could be done using technology such as drones.
Step 4: Lighting management plan
This should include all relevant project information (Step 1) and biological information (Step
2). Maps of important bat areas and/or potential conflict areas should be integrated into the
planning process. The lighting management plan should outline proposed mitigation. For a
range of bat specific mitigation measures please see the Bat Light Mitigation Toolbox below.
The plan should also outline the type and schedule for biological and light monitoring to ensure
mitigation is meeting the objectives of the plan and triggers for revisiting the risk assessment
phase of the EIA. The plan should outline contingency options for additional mitigation or
compensation if biological and light monitoring or compliance audits indicate that mitigation is
not meeting objectives (e.g. artificial light is visible from bat roosts or roost populations
decline).
Step 5: Biological and light monitoring and auditing
The success of the impact mitigation and light management should be confirmed through
monitoring and compliance auditing and the results used to facilitate an adaptive management
approach for continuous improvement and contribute to scientific knowledge information
baselines.
Relevant biological monitoring is described in Step 2 above. Concurrent light monitoring
should be undertaken and interpreted in the context of how bats and their prey perceive light
and within the limitations of monitoring techniques described in Measuring Biologically
Relevant Light. Auditing, as described in the lighting management plan, should be undertaken
to ensure artificial lighting at the site is consistent with the lighting management plan and
relevant conservation objectives.
Step 6: Review
The EIA should incorporate a continuous improvement review process that allows for
upgraded mitigations, changes to procedures and renewal of the lighting management plan
based on biological monitoring of artificial light impacts on bats.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
111
Bat Light Mitigation Toolbox
Appropriate lighting design/lighting controls and mitigating the effect of artificial light will be
site, project and species specific. Where no data is available regarding how artificial light
affects a particular bat species / behaviour / habitat, the precautionary principle should be
followed and light pollution should be reduced. How lighting impacts insects should also be
considered as this is relevant for many bat species.
All projects should incorporate the Best Practice Light Design Principles. Table 19 provides a
toolbox of management options relevant to bats. These should be implemented in addition to
the six Best Practice Light Design principles. Not all mitigation options will be practicable for
every project. Table 20 provides a suggested list of light types appropriate for use near
important bat habitat and those to avoid.
The most effective management actions for mitigating the impact of artificial lights for bats
include:
• maintaining natural darkness for all bat habitat
• maintaining dark, unlit corridors from roosts to foraging areas
• removing or redirecting artificial light directed at roosts or in their immediate vicinity
• removing, redirecting, or shielding artificial lights in known foraging areas keeping
intensity as low as practicable, noting that incident light below 1 lx has been
demonstrated to be disruptive to some bat species (Azam et al., 2018)
Other mitigation measures that may be less effective, but could be considered, include:
• implementing part-night lighting schemes
• modification of luminaires to narrow spectrum, longer wavelengths (such as red light)
(Spoelstra et al., 2017; Haddock, 2018)
• installing motion sensor lighting, noting that this may cause a startle response and
assessment of its effectiveness as a mitigation tool will be needed
Table 19: Light management options for bats
Management Action
Detail
Avoid adding artificial light to previously
unlit areas.
Maintaining dark areas is crucial when
managing effects on nocturnal species such
as bats.
Implement appropriate mitigation where
and when bats are likely to be present.
Roosts, commuting routes, foraging areas
and water sources are areas used by bats
that are most likely to be affected by
artificial light. Any direct or indirect artificial
light that is visible to a person standing in
foraging habitats, commuting corridors or
roost habitats will potentially be visible to a
bat and should be modified to prevent it
being seen.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
112
Management Action
Detail
Turn out lights for as much of the night as
possible.
Exterior lights or interior lights which spill
light outside should be turned off for as
much of the night as possible to prevent
negative impacts on bats.
Keep exterior lighting to a minimum.
Only light where necessary and minimise
intensity. Stay below legally allowed light
levels with outdoor lighting (noting that
often these light levels are not legal, but
example those developed by the IES and
CIE in addition to legal prescriptions). Note
that good visibility for humans depends on
avoiding too high contrasts between max.
and min. visible luminance. If visible
luminance is reduced, e.g. by shielding or
suitable optical design, overall lower
illuminance levels can achieve an even
better visibility then higher illuminance
levels, if the visible luminance is also
higher.
Use motion sensors to turn lights on only
when needed.
This will mean areas remain dark for longer
periods. LEDs have no warm up or cool
down limitations so can remain off until
needed and provide instant light when
required. However, consider whether this
will trigger a startle response for bats.
Avoid high intensity light of any colour.
Keeping light intensity as low as possible
will reduce impacts on bats.
Use lights with reduced or filtered blue
wavelengths.
Bats and their prey are particularly sensitive
to short wavelength light. Blue light
influences the circadian rhythm of
vertebrates and can cause a shift of
sleep/activity patterns.
Avoid violet and ultraviolet wavelengths.
While circadian effects are lower with violet
than with blue wavelengths, insect
attraction can be higher which can have
implications for insectivorous bats.
Use LEDs with warmer spectral
composition (<<2,700 K).
Even if there is no strict correlation of blue
content to CCT, most white light sources
with low CCT, i.e. warmer colour
temperatures, also have lower blue content.
Reviewing the amount of short wavelength
light present in each light type using a
spectral power curve is important to
manage short wavelength light.
Reduce visibility of light sources by
minimising radiance, using shielding and
lowering luminaire height.
Even distant light sources may attract
wildlife because of their high luminance and
visibility from a far distance and so actions
should be taken to minimise radiance.
Do not illuminate important habitats and
features including roosts, roost
entrances/exits, caves, hibernacula,
swarming sites, associated flightpaths,
These important habitats should be kept
dark by avoiding irradiance at these sites.
They should not be illuminated with any

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
113
Management Action
Detail
commuting habitat, foraging areas
(including urban parks, gardens, forest
edges, hedgerows) and drinking sites.
spectra (including red light) because any
light can have negative effects.
Do not illuminate façades of buildings that
are close to important bat habitat
e.g. roosts, caves, hibernacula, swarming
sites, associated flightpaths, commuting
habitat, foraging areas and water sources.
Building façades should not be illuminated
in order to reduce light pollution in general,
but this is particularly important in areas
close to/in bat habitat. Buildings which are
known to house roosts should not be
illuminated during the whole reproductive
season.
Maintain natural light/dark levels (as
measured at the new moon) at roost
entrances, exits and emergence corridors.
Bats are particularly sensitive in these
locations because of the risk of predation
and so natural light/dark levels should be
maintained.
Do not illuminate flyways between roost
entrances/exits and hedgerows, treelines
and other commuting routes.
Lighting can disrupt commuting routes
leading to increased flight time and energy
expenditure. Where feasible natural
light/dark levels (new moon) should be
maintained.
Avoid illumination at foraging areas such as
water bodies (rivers, ponds, canals) and
forests, as well as at drinking sites,
including small ponds and livestock drinking
troughs.
Bats can be deterred from foraging and
drinking sites if they are illuminated and so
these areas should be maintained with
natural light/dark levels (as measured at the
new moon).
Discourage visits to caves with bats
present, particularly those with
nursery/maternity colonies or hibernating
bats so that there is no risk of artificial light
being introduced e.g. via flashlights /
torches, or more permanent lighting.
Some areas are only used by bats
seasonally and light management should
take this into consideration.
Minimise lighting and its duration in caves.
Only use lights when needed and limit them
to areas away from bats.
Ideally do not light caves where bats are
present. If necessary, only illuminate
specific cave formations rather than the
whole cave. Switch off lights when not
needed.
Seek to separate lights, including
streetlights, from important bat habitats by
an appropriate distance, and using
shielding and other measures to reduce
light spill where appropriate.
Distance alone may not be enough. Good
optical quality of luminaires is required to
prevent spill light to locations away from the
street. Shielding and other measures to
reduce light spill should be implemented.
Avoid directing light onto vegetation/plants.
Insectivorous bats may forage near
vegetation and nectar/fruit feeding bats
feed directly from plants and therefore light
on vegetation should be avoided.
If lights need to be installed inside buildings
with roosts, use low intensity and highly
directed light sources away from the bats.
Use light only temporarily and when
needed.
Light should only illuminate the direct
pathways of humans when needed to
ensure their safety and should be switched
off when not necessary. Automatic timers
can be used. Lights should be automatically
turned off when it gets dark so that lights
are not accidentally left on throughout the
night.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
114
Management Action
Detail
Install lights at lower heights so only target
areas are illuminated, for example in
underpasses or by using bollard lights to
light paths.
Lights installed at lower heights will help
reduce light spill and unnecessary lighting
of dark areas.
Use other materials such as glow in the
dark or light-coloured paths.
In some circumstances lighting may not be
necessary for human orientation if
alternative materials are used to highlight
paths or to mark critical objects e.g. curbs
or paths.
Create buffer zones between key bat
habitat and areas to be lighted.
The key habitat should be maintained with
no artificial light, the area next to the key
habitat should have strictly limited
illuminance, the area next to that should be
moderately illuminated with the use of light
barriers or screening, and, in the main
development area where lighting is deemed
most necessary, illuminance levels should
be kept as low as possible. See Bat
Conservation Trust and ILP (2018) for a
useful diagram illustrating this.
Use non-reflective, dark-surfaced buildings,
walls, fences, and soft landscaping to block
light spill where appropriate. Vegetation
may also be used as a buffer.
Though it is preferable to avoid light spill by
installing high quality luminaires, if this is
not enough, residual spill light can further
be reduced by blocking it with walls, fences,
soft landscaping or additional shielding.
Where vegetation is used as a buffer,
ensure that it is not directly illuminated.
Use orientation of light to mitigate negative
impacts.
Light should never be directed towards
habitats, drinking zones or other critical
areas where bats are present. Adapting the
orientation of luminaires can help to
minimise spill light towards critical/key
areas.
Consider placement of footpaths, open
space, and number/size of windows in new
developments to minimise light spill on to
key habitat.
The location of areas and pathways which
need to be illuminated should be oriented
away from habitats to reduce impacts.
Install dimmable streetlights in areas
where roads cross important bat habitats.
Dim lights to lowest allowable levels.
Streetlights can be dimmed depending on
time of day, to reduce light levels in critical
times for bats (e.g. 2 hours after sunset),
but can also be dimmed depending on
traffic, so that they only turn on if traffic is
detected. LED streetlights have no delay to
ramp up light levels within seconds.
Only light areas at times when the light is
necessary. Ideally, start the dark phase of a
lighting scheme within the first two hours
after sunset to reduce impacts.
The first two hours after sunset are most
critical for disturbance of bats by artificial
light because this often overlaps with times
bats emerge from their roosts and are most
active. Timing of lighting schemes should
take this into account and ensure darkness
or extreme low light levels during this time.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
115
Management Action
Detail
Use motion sensors and timers to reduce
lighting periods to when lighting is
necessary.
The trigger threshold should be set high
(so that only large objects like humans
trigger the sensors) and the trigger
duration should be appropriately short (no
more than a few minutes). Note that these
devices require some degree of attention
and maintenance.
Control lighting when bats are present and
consider seasonal activities of bats,
including migration, mating, and raising
dependent young, to make appropriate
lighting choices.
For example, in buildings which are used by
bats only for a short period of time during
the year, external lighting towards these
buildings should be completely avoided
during the period that bats are present.
Table 20 Commercial luminaire types that are considered generally less impactful for use near
bat habitat, and those to avoid
Light type
Suitability for use near bat habitat
Low-pressure sodium vapour
High-pressure sodium vapour
Filtered LED *
Filtered metal halide *
Filtered white LED *
Narrowband Amber LED
PC Amber
White LED
Metal halide
White fluorescent
Halogen
* only if a filter approved by the manufacturer is applied to remove
the short-wavelength (< 500 nm) light.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
116
Glossary
ACAP is the Agreement on the Conservation of Albatrosses and Petrels.
ALAN is Artificial Light At Night and refers to artificial light outside that is visible at night.
Artificial light is composed of visible light as well as some ultraviolet (UV) and infrared (IR)
radiation that is derived from an anthropogenic source.
Artificial skyglow is the part of the sky glow that is attributable to human-made sources of
light (see also skyglow).
Biologically relevant is an approach, interpretation or outcome that considers either the
species to which it refers, or factors in biological considerations in its approach.
Brightness is the strength of the visual sensation on the naked eye when lit surfaces are
viewed.
Bulb is originally a traditional source of electric light and is a component of a luminaire. Bulbs
are also available as LED-retrofits with the same geometry as traditional bulbs for retrofitting
in old luminaires. Modern LED luminaires do not have bulbs as light sources, but the LED are
mounted on electronic boards.
Candela (cd) (photometric term) is a basic photometric unit of illumination that measures
the amount of light emitted in the range of a (three-dimensional) angular span, corresponding
to the luminous flux per solid angle in lm/sr. This should not be confused with the unit for
luminance which is typically measured in candela per square metre (cd/m
2
) and includes the
area of the light source.
Charge Coupled Device (CCD) is the sensor technology used in digital cameras. It converts
captured light into digital data (images) which can be processed to produce quantifiable data.
CIE
sets most international lighting standards. The most relevant international lighting standards
are first published by CIE and later as a joint standard by CIE and ISO.
CMS is the Convention on the Conservation of Migratory Species of Wild Animals or the Bonn
Convention.
Colour temperature is used to describe the perceived colour of a white light source ranging
from cold white (bluish) to warm white (yellowish), measured in Kelvin (K). Colour temperature
is only used for black-body radiators and is in this case corresponding to their real
temperature,500 K will have a warm
appearance while 6,500 K will appear cold.
Commuting routes are flight paths that are used regularly by bats to fly from a roost to a
foraging area (and back) or to move between foraging areas or between roosts.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
117
Correlated Colour Temperature (CCT) is used to characterise the perceived colour of an
artificial white light source. It is correlated to the response of the human eye. Correlated colour
temperature is expressed in Kelvin (K).
Cumulative light refers to increased sky brightness due to light emissions contributions from
multiple light producers. Measured as skyglow.
Disorientation refers to any species moving in a confused manner e.g. a turtle hatchling
circling and unable to find the ocean.
EIA is an Environmental Impact Assessment process.
Electromagnetic radiation is a kind of radiation including visible light, radio waves, gamma
rays, and X-rays, in which electric and magnetic fields vary simultaneously.
Fallout refers to birds that collide with structures when disoriented.
Feeding buzzes are stereotypic sequences of echolocation calls indicating feeding is taking
place.
Footcandle (fc or ftc) (photometric term) is a unit of illuminance used in America, it is based
on the brightness of one candle at a distance of one foot. Measured in lumens per square foot,
one ftc is equal to approximately 10.7639 lux. This is not an appropriate measure for
understanding how animals perceive light. It should not be used in international documents,
as it is not compliant to the international system of units SI.
FMP refers to the Field Management Program.
Genetic population (also known as genetic stock) is a discrete grouping of a species by
genetic relatedness. Management of the species may be undertaken on a genetic population
basis because each genetic population represents a unique evolutionary history which, if lost,
cannot be replaced.
Glare refers to a condition of reduced or disabled visibility due to high luminance or extreme
luminance contrasts. As glare is related to disturbing a visual task in humans, when the
luminaire is properly mounted for its application, "low glare" luminaires may nevertheless
exhibit very high visible luminance depending on the viewing angle under which the light
source appears.
Grounding refers to events where birds fail to take their first flight from the nest or collide with
a structure (adults and juveniles) and are unable to launch back into the air.
Hibernacula.
Horizontal plane, in relation to the light fitting, means the horizontal plane passing through
the centre of the light source (for example the bulb) of the light fitting.
HPS is a high-pressure sodium lamp that produces a characteristic wavelength near 589 nm.
Illuminance is a photometric measure of the total luminous flux incident on a surface, per
unit area. It is a measure of how much the incident light illuminates the surface, wavelength-
weighted to correlate with human brightness perception. Illuminance is measured in lux (lx)
or equivalently in lumens per square metre (lm/m
2
).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
118
Important habitats are those areas that are necessary for an ecologically significant
proportion of a species to undertake important activities such as foraging, breeding, roosting
or dispersal. Important habitats will be species specific and will depend on their conservation
status.
Incandescent bulb is a bulb that provides light by a filament heated to a high temperature by
electric current. Its sale is banned in most countries because of its low energy efficiency.
Intensity is the amount of energy or light in a given dire
can be used as a surrogate for illuminance or luminance, irradiance and all qualities related to
light. Intensity per se is not a defined lighting term and should be avoided as soon as specific
quantities (including units) need to be used or if specific effects of light are discussed. It can
be used in a descriptive way, but not as a formal quantity.
IR is infrared radiation and represents a band of the electromagnetic spectrum with
wavelength from 780 nm to 1 mm.
Irradiance (radiometric term) is a measurement of radiant flux at or on a known surface area,
in W/m
2
. This measure is more appropriate for understanding animal perception of light but
needs to be weighted with the spectral sensitivity of a specific animal for the wavelengths
contained in the perceived radiation.
ISO is the International Organization for Standardization. The fundamental CIE standards on
light and lighting are also published as ISO standards.
IUCN is the International Union for the Conservation of Nature.
Kelvin (K) is the absolute unit for temperature and is equal in magnitude to one degree Celsius
(° C), but with a different zero point (0 °C = 273 K). Kelvin is typically used to describe
Correlated Colour Temperature (CCT). 6,000 K corresponds to the colour impression of a
black body radiator at a surface temperature of 5,727 °C.
Lamp
Examples include incandescent, fluorescent, high-intensity discharge (HID) lamps, and low-
pressure sodium (LPS) lamps, as well as light-emitting diode (LED) modules and arrays. In
modern LED luminaires, the LED are mounted on electronic boards which are denominated
arrangements integrated in the
traditional shapes of former classical light sources.
LED is a light-emitting diode, or a semiconductor light source that emits light when current
flows through it. This process works mainly for blue, red and green LED. For white LED, see
Phosphor-converted LED (PC-LED).
Light fitting (luminaire) is the complete lighting unit. It includes the bulb, reflector (mirror) or
refractor (lens), the ballast, housing and the attached parts.
Light is the radiant energy that is visible to humans. Light stimulates receptors in the visual
system and those signals are interpreted by the brain making things visible. As animals have
different sensitivities for vision, wavelengths, which are not considered as light, can be
perceived by animals. Such wavelengths are denominated as radiation.
Light pollution refers to artificial light that alters the natural patterns of light and dark in
ecosystems.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
119
Light spill is the light that falls outside the boundaries of the object or area intended to be lit.
Spill light serves no purpose and if directed above the horizontal plane, contributes directly to
artificial skyglow. Also called spill light, obtrusive light or light trespass.
Lighting controls are devices used for either turning lights on and off, or for dimming.
LNG is liquefied natural gas.
LPS is a low-pressure sodium lamp that produces a characteristic wavelength near 589 nm.
Lumen (lm) (photometric term) is the unit of luminous flux, a measure of the total quantity
of visible light emitted by a source per unit of time. This is a photometric unit, weighted to the
sensitivity of the human eye. If a light source emits one candela of luminous intensity uniformly
across a solid angle of one steradian, the total luminous flux emitted into that angle is one
lumen. A point light source having a homogeneous luminous intensity of one candela in any
direction, emits a total luminous flux of 12.57 lm.
Luminaire refers to the complete lighting unit (fixture or light fitting), consisting of a lamp, or
lamps and ballast(s) (when applicable), together with the parts designed to distribute the light
(reflector, lens, diffuser), to position and protect the lamps, and to connect the lamps to the
power supply.
Luminous flux is the total light emitted by a bulb in all directions which is measured in lumen.
Luminance (cd/m
2
) is a photometric measure of the luminous intensity per unit area of light
travelling in a given direction, wavelength-weighted to correlate with human brightness
perception. Luminance is measured in candela per square metre (cd/m
2
). Luminance and
illuminance ("Lux") are related, in the sense that luminance is a measure of light emitted from
a surface (either because of reflection or because it is a light-emitting surface) in a certain
direction, and illuminance is a measure for light hitting a surface.
Lux (lx) is a photometric unit for the level of illumination of a surface. The difference between
lux and candela is that lux measures the illumination of a surface as luminous flux per area
(in lm/m
2
), while candela is the unit for the quantity of light emitted in a certain solid, angle.
Both units are based on human sensitivity and are not an appropriate measure for
understanding how animals perceive light.
Magnitudes per square arc second (magnitudes/arcsec
2
) (radiometric term) is a term
used in astronomy to measure sky brightness within an area of the sky that has an angular
area of one second by one second. The term magnitudes per square arc second means that
the brightness in magnitudes is spread out over a square arcsecond of the sky. Each
magnitude lower (numerically) means just over 2.5 times more light is coming from a given
patch of sky. A change of 5 magnitudes/arcsec
2
means the sky is 100x brighter.
Misorientation occurs when a species moves in the wrong direction, e.g. when a turtle
hatchling moves toward a light and away from the ocean.
Mounting height is the height of the fitting or bulb above the ground.
Nanometer (nm) is the unit used for wavelength. 1 nm = 10
-9
m. = 1 billionths of a metre or 1
millionth of a millimetre. It is used as the unit for the wavelength of optical radiation.
Wavelengths larger than 1,000 nm, e.g for infrared radiation, are described in µm (micro
meter). 1 µm = 1,000 nm.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
120
Natural skyglow is that part of the skyglow that is attributable to radiation from celestial
Phosphor Converted LED (PC-LED). The LED chip (semiconductor) produces blue or violet
light which is partially converted to different colours by a phosphor layer, which covers the
LED chip. The phosphor emits visible light with longer wavelengths than the absorbed blue or
violet light and the light emitted from the LED surface is a mixture of the light from the phosphor
and the residual light from the LED-chip. Standard phosphors are mixtures of different crystals,
and all white LED are PC-LED. The Correlated Colour Temperature (CCT) is determined by
the mixture and the thickness of the phosphor. In addition to white light with different CCT,
new phosphor mixtures are allowing LED with amber, red or other coloured light emission to
be built. The spectral width from a phosphor emission typically covers a wider spectral range
than the emission from only the LED-semiconductor.
Photometric terms refer to measurements of light that are weighted to the sensitivity of the
human eye. They do not include the shortest or the longest wavelengths of the visible
spectrum for animals and so are not appropriate for understanding the full extent of how
animals perceive light.
Photometry is a subset of radiometry that is the measurement of light as it is weighted to the
sensitivity of the human eye.
Photoperiod refers to the daylight fraction of the 24 hour day, which changes across the year
except at the equator. Photoperiod can be manipulated by artificial light.
Photopic vision refers to human vision under well-lit conditions. It allows colour perception,
in contrast to scotopic vision at low light levels, which allows us to see only on a blue-grey
impression.
Phototaxis is the tendency of an organism to move in a certain direction depending on the
light distribution at its place. This is equivalent to orientation on the direction of light incident.
Positive phototaxis means that movement goes towards increased brightness, resulting in
attraction by light. Negative phototaxis is also possible, resulting in avoidance of light.
Point source is a light source which emits light from a small area usually in all directions. LED
point sources emit in a hemisphere. Without shielding point sources allow to be seen directly
and exhibit the risk of strong glare.
Radiance (radiometric term) is a measure for density of radiant intensity with respect to
projected area in a specified direction at a specified point, measured in W/(m
2
sr)
Radiant flux/power (radiometric term) is expressed in watts (W). It is the total optical
power of a light source. It is the radiant energy emitted, reflected, transmitted or received,
per unit time. Sometimes called radiant power, and it can also be defined as the rate of flow
of radiant energy.
Radiant intensity (radiometric term) is the density of radiant flux (power) emitted in a known
solid angle, W/steradian, and has a directional quantity.
Radiometric terms refer to light measured across the entire optical spectrum (not weighted
to the human eye). These are appropriate for understanding how animals perceive light.
Radiometry is the measurement of all wavelengths across the entire optical spectrum (not
weighted to the human eye).

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
121
Reflected light is light that bounces off a surface. Light coloured surfaces reflect more light
than darker coloured surfaces.
RGB stands for Red, Green and Blue. These are the colours which the human eye is sensitive
to. Red, Green and Blue light sources can be used to mix other colours visible for humans. In
digital cameras the light is separated in these three primary colours and measured separately.
Colour images consist of three layers of the same image, one each for blue, green and red.
Roosts are locations used by bats at different times for different activities. Depending on the
species, roosts may be in buildings, barns, caves, mines, trees, tree hollows, etc. Different
types of roosts are listed below:
Day roost A place where bats rest or shelter in the day. Night roost A place where
bats rest or shelter during the night. May be used by a single individual or by an entire
colony. Night roosts may also be used as day roosts. Feeding roost A place where
bats rest or feed during the night. Transitional/Occasional roost Used by a few
individuals or occasionally small groups for generally short periods of time. Maternity
roost Where female bats give birth and/or raise their young. Hibernation roost
Where bats may be found individually or together during winter. They often have a
constant cool temperature and high humidity.
Scotopic vision refers to vision during low-light or almost dark conditions, related to human
sensitivity. Other species may see well under scotopic conditions.
Sensitive receptor is any living organism that has increased sensitivity or exposure to
environmental contaminants that may have adverse effects.
Sensor is an electronic device used in lighting to turn light on or off or to dim or brighten it.
Presence sensors are used to detect the presence of humans or objects (e.g. vehicles) with
the intention to dim down or switch off the light, when no presence is detected. Light sensors
measure available natural or ambient light and dim down or switch off artificial light, if natural
light levels are sufficient. They can also ensure that artificial light is only added in the needed
quantity to achieve a certain target level of light (e.g. at workplaces). Use of sensors saves
energy and prevents the application of light when it is not needed.
Shielded light fitting is a physical barrier used to limit or modify the light paths from a
luminaire.
Skyglow is the brightness of the night sky caused by the cumulative impact of reflected
radiation (usually visible light), scattered from the constituents of the atmosphere in the
direction of observation. Skyglow comprises two separate components: natural skyglow and
artificial skyglow (see also natural skyglow and artificial skyglow).
Smart controls are devices to vary the intensity or duration of operation of lighting, such as
motion sensors, light sensors, timers and dimmers used in concert with outdoor lighting
equipment.
Spectral power distribution provides a representation of the spectral power emitted from a
light source at each wavelength. It can be visualised in a graph as a curve of intensity vs.
wavelength or in a table.
Swarming is a behaviour exhibited by some bat species.
of some temperate bat species that occurs from late summer to autumn. Plecotus auritus

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
122
before dawn. Swarming is an important part of social interactions, including courtship. Some
entrance to a roost (especially maternity roosts) before the bats enter at dawn.
Task lighting is used to provide direct light for specific activities without illuminating the entire
area or object.
Upward Light Ratio (ULR) or Upwards Light Output Ratio (ULOR) is the proportion of the
light (flux) emitted from a luminaire or installation that is emitted at and above the horizontal,
excluding reflected light when the luminaire is mounted in its parallel position. ULR is the
upward flux/total flux from the luminaire.
UV (Ultraviolet radiation) is electromagnetic radiation with wavelengths from 400 nm to 100
nm, shorter than that of visible light but longer than X-rays. UV is not visible for humans, but
can be visible for many nocturnal animals and insects.
Visible light transmittance (VLT) is the proportion of light transmitted by window glass which
is recorded as either TVw (visible transmittance of the window) and is reported as a
dimensionless value between 0 and 1, or 0 and 100%. A low TVw (e.g. < 30%) indicates little
light is transmitted through the glass while higher TVw values are associated with increasing
light transmittance. While the VLT/Tvw rating varies between 0 and 1, most double-glazed
windows rate between 0.3 and 0.7, which means that between 30% and 70% of the available
light passes through the window.
W/m
2
is a measure of irradiance, the radiant power irradiated on a unit area of a surface. This
is an appropriate measure for understanding how animals perceive light, when weighted with
ty for the radiation.
Wattage is the electrical power needed to light a light source. Generally, the higher the
wattage, the more lumens are produced with the same type of light source. LED can produce
more lumens with lower wattage than traditional light sources. Higher wattage and more
lumens give a brighter light.
Wavelength is a physical property attributed to the energy of a photon. Short wavelengths
photons have higher energy than longer wavelengths photons. Spectral power distributions of
light sources show the intensity (corresponding to the number of photons) at specific
wavelengths. For the visible part of radiation, the wavelength is also correlated to the colour
impression. Ultraviolet and blue light are examples of short wavelength light while red and
infrared light is long wavelength light. The wavelength of optical radiation is measured in
nanometers (humans can see radiation between 380 nm and 780 nm).
Zenith is an imaginary point directly above a location, on the imaginary celestial sphere.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
123
References
Adams CA, Fernández-Juricic E, Bayne EM, et al. (2021) Effects of artificial light on bird
movement and distribution: a systematic map. Environmental Evidence 10:37.
Ainley DG, Podolsky R, Nur N, et al.
shearwater on Kauai: a model for threatened petrels on urbanized tropical oceanic islands.
Studies in Avian Biology 22:108-123.
Algvere PV, Marshall J and Seregard S (2006) Age‐related maculopathy and the impact of
blue light hazard. Acta Ophthalmologica Scandinavica 84(1):4-15.
Allen JA (1880) Destruction of birds by light-houses. Bulletin of the Nuttall Ornithological
Club 5:131-138.
American Bird Conservancy (2023) Red Knot. Available at: https://abcbirds.org/bird/Red-
Knot/
Angers K, Haddad N, Selmaoui B, et al. (2003) Effect of melatonin on total food intake and
macronutrient choice in rats. Physiology and Behavior 80:9-18.
Atchoi E, Mitkus M and Rodríguez A (2020) Is seabird light-induced mortality explained by
the visual system development? Conservation Science and Practice 2: e195.
Austad M, Oppel S, Crymble J, et al. (2023) The effects of temporally distinct light pollution
from ships on nocturnal colony attendance in a threatened seabird. Journal of Ornithology
https://doi.org/10.1007/s10336-023-02045-z
Azam C, Le Viol I, Bas Y, et al. (2018) Evidence for distance and illuminance thresholds in
the effects of artificial lighting on bat activity. Landscape and Urban Planning 175: 123-135.
Aziz SA, McConkey KR, Tanalgo K, et al. (2021) The critical importance of Old World fruit
bats for healthy ecosystems and economies. Frontiers in Ecology and Evolution 9: 641411.
Bamford M, Watkins D, Bancroft W, et al. (2008) Migratory Shorebirds of the East Asian-
Australasian Flyway; Population Estimates and Internationally Important Sites: Wetlands
International - Oceania: Canberra, Australia. 249.
Barentine JC (2019) Methods for Assessment and Monitoring of Light Pollution around
Ecologically Sensitive Sites. Journal of Imaging 5(54).
Barghini A and Souza de Medeiros BA (2012) UV radiation as an attractor for insects.
Leukos 9(1): 47-56.
Barré K, Kerbiriou C, Ing R-K, et al. (2021) Bats seek refuge in cluttered environment when
exposed to white and red lights at night. Movement Ecology 9: 3.
Bat Conservation Trust and Institution of Lighting Professionals (ILP) (2018) Guidance Note
08/18. Bats and artificial lighting in the UK. Available at:
https://cdn.bats.org.uk/uploads/pdf/Resources/ilp-guidance-note-8-bats-and-artificial-lighting-
compressed.pdf?v=1542109349

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
124
Bat Conservation Trust (2023a) Foraging habitats. Available at:
https://www.bats.org.uk/about-bats/where-do-bats-live/bat-habitats/foraging-habitats
Bat Conservation Trust (2023b) Bat habitats. Available at: https://www.bats.org.uk/about-
bats/where-do-bats-live/bat-habitats/commuting-habitats
Battersby J (comp.) (2017) Guidelines for Surveillance and Monitoring of European Bats.
EUROBATS Publication Series No. 5. UNEP/EUROBATS Secretariat, Bonn, Germany, 95
pp. 3
rd
edition. Available at:
https://www.eurobats.org/sites/default/files/documents/publications/publication_series/EURO
BATS_PublSer_No5_3rd_edition.pdf
Battley PF, Warnock N, Tibbitts TL, et al. (2012) Contrasting extreme long-distance
migration patterns in bar-tailed godwits Limosa lapponica. Journal of Avian Biology 43(1):21-
32.
Beadnell CM (1937) The Toll of Animal Life exacted by Modern Civilisation. Proceedings of
the Zoological Society of London A107(2): 173-182.
Benenson W, Harris JW, Stcker H, et al. (eds.) (2006) Handbook of physics. Springer
Science and Business Media.
Bennie J, Davies TW, Cruse D, et al. (2016) Ecological effects of artificial light at night on
wild plants. Journal of Ecology 104(3):611-620.
Berson DM (2007) Phototransduction in ganglion-cell photoreceptors. Pflgers Archiv
454(5):849-855.
Bird BL, Branch LC and Miller DL (2004) Effects of coastal lighting on foraging behaviour on
beach mice. Conservation Biology 18:1435-1439.
Black A (2005) Light induced seabird mortality on vessels operating in the Southern Ocean:
incidents and mitigation measures. Antarctic Science 17:67-68.
Boldogh S, Dobrosi D and Samu P (2007) The effects of the illumination of buildings on
house-dwelling bats and its conservation consequences. Acta Chiropterologica. 9(2): 527-
534.
Bolliger J, Hennet T, Wermelinger B, et al. (2020) Effects of traffic-regulated street lighting
on nocturnal insect abundance and bat activity. Basic and Applied Ecology 47: 44-56.
Bolton D, Mayer-Pinto M, Clark GF, et al. (2017) Coastal urban lighting has ecological
consequences for multiple trophic levels under the sea. Science of the Total Environment
576:1-9.
Bowmaker JK and Martin GR (1985) Visual pigments and oil droplets in the penguin,
Spheniscus humboldti. Journal of Comparative Physiology A 156(1):71-77.
Bowmaker JK, Heath LA, Wilkie SE, et al. (1997) Visual pigments and oil droplets from six
classes of photoreceptor in the retinas of birds. Vision Research 37:2183-2194.
Boyes DH, Evans DM, Fox R, et al. (2021) Is light pollution driving moth population
declines? A review of causal mechanisms across the life cycle. Insect Conservation and
Diversity 14(2): 167-187.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
125
Briedis M, Bauer S, Adamík P, et al. (2019) Broad-scale patterns of the Afro-Palaearctic
landbird migration. Global Ecology and Biogeography 29(4): 722-735.
Bruderer B, Peter D and Steuri T (1999) Behaviour of migrating birds exposed to X-band
radar and a bright light beam. Journal of Experimental Biology 202(9): 1015-1022.
Bruderer B, Peter D and Korner-Nievergelt F (2018) Vertical distribution of bird migration
between the Baltic Sea and the Sahara. Journal of Ornithology 159: 315-336.
Burgin CJ, Wilson DE, Mittermeier RA, et al. (2020) Illustrated Checklist of the Mammals of
the World. Volume 2. Eulipotyphla to Carnivora. Lynx editions, Barcelona.
Cabrera-Cruz SA, Smolinsky JA and Buler JJ (2018) Light pollution is greatest within
migration passage areas for nocturnally-migrating birds around the world. Nature Scientific
Reports 8: e3261.
Cabrera-Cruz SA, Cohen EB, Smolinsky JA, et al. (2020) Artificial Light at Night is Related to
Broad-Scale Stopover Distributions of Nocturnally Migrating Landbirds along the Yucatan
Peninsula, Mexico. Remote Sensing 12: 395.
Cabrera-Cruz SA, Larkin RP, Gimpel ME, et al. (2021) Potential Effect of Low-Rise,
Downcast Artificial Lights on Nocturnally Migrating Land Birds. Integrative and Comparative
Biology 61(3): 1216-1236.
Campbell C (1994) The effects of flash photography on nesting behavior of green turtles
(Chelonia mydas) at Tortuguero, Costa Rica. In Proceeding of the fourteenth annual
symposium on sea turtle biology and conservation. 1994. NOAA Technical Memorandum -
NMFS-SEFSC.
Campbell AL, Naik RR, Sowards L, et al. (2002) Biological infrared imaging and sensing.
Micron 33(2): 211-225.
Campos SMC (2017) The impact of artificial lighting on nature. Paper presented at 6th
SENAC MEETING of Integrated Knowledge, São Paulo, 18 May 2017.
Cannell BL and Cullen JM (1998) The foraging behaviour of little penguins Eudyptula minor
at different light levels. Ibis 140(3):467-471.
Capuska GEM, Huynen L, Lambert D, et al. (2011) UVS is rare in seabirds. Vision research
51(12):1333-1337.
Chevron Australia (2018) Gorgon Gas Development and Jansz Feed Gas Pipeline Long-
term Marine Turtle Management Plan. 83.
Cianchetti-Benedetti M, Becciu P, Massa B, et al. (2018) Conflicts between touristic
recreational activities and breeding shearwaters: short-term effect of artificial light and sound
on chick weight. European Journal of Wildlife Research 64:19.
CIE 115:2010: Lighting of Roads for Motor and Pedestrian Traffic. Available at:
https://cie.co.at/publications/lighting-roads-motor-and-pedestrian-traffic-2nd-edition
CIE 150:2017 Guide on the Limitation of the Effects of Obtrusive Light. Available at:
https://cie.co.at/publications/guide-limitation-effects-obtrusive-light-outdoor-lighting-
installations-2nd-edition

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
126
CIE 017:2020 ILV: International Lighting Vocabulary, 2
nd
edition. Available at:
https://cie.co.at/e-ilv
CMS (2023a) Species. Available at: https://www.cms.int/en/species
CMS (2023b) Central Asian Flyway. Available at:
https://www.cms.int/en/legalinstrument/central-asian-flyway
Cochran WW and Graber RR (1958) Attraction of nocturnal migrants by lights on a television
tower. The Wilson Bulletin, 70(4), pp.378-380.
Cohen EB, Horton KG, Marra PP, et al. (2021) A place to land: spatiotemporal drivers of
stopover habitat use by migrating birds. Ecology Letters 24(1): 38-49.
Colling OM, Guglielmo CG, Bonner SJ, et al (2022) Migratory songbirds and urban window
collision mortality: Vulnerability depends on species, diel timing of migration, and age
class. Avian Conservation and Ecology, 17(1), p.22.
Collins J (ed.) (2016) Bat Surveys for Professional Ecologists: Good Practice Guidelines (3
rd
edition). The Bat Conservation Trust, London. Available at:
https://www.bats.org.uk/resources/guidance-for-professionals/bat-surveys-for-professional-
ecologists-good-practice-guidelines-3rd-edition
Colwell MA (2010) Shorebird ecology, conservation, and management. Berkeley, California:
University of California Press. p. 344
Commonwealth of Australia (2016) National Recovery Plan for the Mountain Pygmy-
possum Burramys parvus Prepared by the Victorian Department of Environment, Land,
Water and Planning: Canberra, Australia. 43.
Conklin JR and Colwell MA (2007) Diurnal and nocturnal roost site fidelity of Dunlin (Calidris
alpina pacifica) at Humboldt Bay, California. The Auk 124(2):677-689.
COPUOS (Committee on the Peaceful Uses of Outer Space) (2021) Recommendations to
Keep Dark and Quiet Skies for Science and Society. Available at:
https://www.iau.org/static/publications/uncopuos-stsc-crp-8jan2021.pdf
Cravens ZM and Boyles JG (2019) Illuminating the physiological implications of artificial light
on an insectivorous bat community. Oecologia 189: 69-77.
Cresswell W (1994) Flocking is an effective anti-predation strategy in redshanks, Tringa
tetanus. Animal Behaviour 47(2):433-442.
Cruz LM, Shillinger GL, Robinson NJ, et al. (2018) Effect of light intensity and wavelength on
the in-water orientation of olive ridley turtle hatchlings. Journal of Experimental Marine
Biology and Ecology 505: 52-56.
Cryan PM, Gorresen PM, Straw BR, et al. (2022) Influencing activity of bats by dimly lighting
wind turbine surfaces with ultraviolet light. Animals 12:9.
Davies TW and Smyth T (2017) Why artificial light at night should be a focus for global
change research in the 21
st
century. Global Change Biology 24: 872-882.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
127
de Jong M, Ouyang JQ, Da Silva A, et al. (2015) Effects of nocturnal illumination on life-
history decisions and fitness in two wild songbird species. Philosophical Transactions of the
Royal Society of London. Series B, Biological Sciences 370:2014012820140128.
den Outer P, Lolkema D, Haaima M, et al. (2011) Intercomparisons of Nine Sky Brightness
Detectors. Sensors 11(10):9603.
Department of Conservation and Fisheries New Zealand (2023) Mitigation Standards to
Reduce Light-induced Vessel Strikes of Seabirds with New Zealand Commercial Fishing
Vessels. Available at: https://www.mpi.govt.nz/dmsdocument/56320-Mitigation-Standards-to-
Reduce-Light-induced-Vessel-Strikes-of-Seabirds-with-New-Zealand-Commercial-Fishing-
Vessels
Deppe L, Rowley O, Rowe LK, et al.
shearwaters (Puffinus huttoni) associated with artificial lighting. Notornis 64(4):181-191.
Desouhant E, Gomes E, Mondy N, et al. (2019) Mechanistic, ecological, and evolutionary
consequences of artificial light at night for insects: review and prospective. Entomologia
Experimentalis et Applicata 167: 37-58.
Dias MP, Granadeiro JP, Lecoq M, et al. (2006) Distance to high- tide roosts constrains the
use of foraging areas by dunlins: Implications for the management of estuarine wetlands.
Biological Conservation 131:446-452.
Dias MP, Martin R, Pearmain EJ et al. (2019) Threats to seabirds: a global assessment.
Biological Conservation 237: 525-537. https://doi.org/10.1016/j.biocon.2019.06.033
Dokter AM, Farnsworth A, Fink D, et al. (2018) Seasonal abundance and survival of North
Nature ecology and
evolution, 2(10), pp.1603-1609.
Dominoni DM, Goymann W, Helm B, et al. (2013) Urban-like night illumination reduces
melatonin release in European blackbirds (Turdus merula): implications of city life for
biological time-keeping of songbirds. Frontiers in Zoology 10: 60.
Dominoni DM and Partecke J (2015) Does light pollution alter daylength? A test using light
loggers on free-ranging European blackbirds (Turdus merula). Philosophical Transactions of
the Royal Society B: Biological Sciences 370(1667): 20140118.
Duriscoe DM (2013) Measuring anthropogenic skyglow using a natural sky brightness
model. Publications of the astronomical society of the Pacific 125:1370-1382.
Dwyer RG, Bearhop S, Campbell HA, et al. (2013) Shedding light on light: benefits of
anthropogenic illumination to a nocturnally foraging shorebird. Journal of Animal Ecology
82:478-485.
Dzul-Cauich HF and Munguía-Rosas MA (2022) Negative effects of light pollution on
pollinator visits are outweighed by positive effects on the reproductive success of a bat-
pollinated tree. The Science of Nature 109(1), 1-11.
Ecker JL, Dumitrescu ON, Wong KY, et al. (2010) Melanopsin-Expressing Retinal Ganglion-
Cell Photoreceptors: Cellular Diversity and Role in Pattern Vision. Neuron 67(1):49-60.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
128
Eckert KL, Bjorndal KA, Abreu-Grobois FA, et al. (eds.) (1999) Research and Management
Techniques for the Conservation of Sea Turtles. IUCN/SSC Marine Turtle Specialist Group
Publication No. 4.: Washington, DC. 235.
EEAFP (2018) EAAFP MOP10/D1: East Asian-Australasian Flyway Partnership 2019-2028
Strategic Plan. Available at: https://www.eaaflyway.net/wp-
content/uploads/2019/07/MOP10_D01_Strategic-Plan-2019-2028_r_MJ.pdf
Eisenbeis G (2006) Artificial night lighting and insects: attraction of insects to streetlamps in
a rural setting in Germany. Ecological Consequences of Artificial Night Lighting (ed. by C.
Rich and T. Longcore), pp. 281304. Island Press, Washington, District of Columbia.
Elmore JA, Hager SB, Cosentino BJ, et al. (2021a) Correlates of bird collisions with buildings
across three North American countries. Conservation Biology, 35(2), pp.654-665.
Elmore JA, Riding CS, Horton KG, et al. (2021b) Predicting bird-window collisions with
weather radar. Journal of Applied Ecology 58(8): 1593-1601.
Erb V and Wyneken J (2019) Nest-to-Surf Mortality of Loggerhead Sea Turtle (Caretta
caretta. Frontiers in Marine Science 6(271):doi:
10.3389/fmars.2019.00271.
Evans Ogden LJ (1996) Collision course: the hazards of lighted structures and windows to
migrating birds. Fatal Light Awareness Program (FLAP), p.3.
Evans WR, Akashi Y, Altman NS, et al. (2007) Response of night-migrating songbirds in
cloud to colored and flashing light. North American Birds 60(4): 476-488.
Evans WR (2010) Response by William R. Evans to: Green light for nocturnally migrating
birds. Ecology and Society 15(3): r1. Available at:
http://www.ecologyandsociety.org/vol15/iss3/resp1/
Faaborg J, Holmes RT, Anders AD (2010) Recent advances in understanding migration
systems of New World land birds. Ecological Monographs 80(1): 3-48.
Falchi F, Cinzano P, Duriscoe D, et al. (2016) The new world atlas of artificial night sky
brightness. Science Advances 2(6):e1600377.
Falcón J, Torriglia A, Attia D, et al. (2020) Exposure to Artificial Light at Night and the
Consequences for Flora, Fauna, and Ecosystems. Frontiers in Neuroscience 14: 602796.
Farnsworth A, Van Doren BM, Hochachka WM, et al. (2016) A characterization of autumn
nocturnal migration detected by weather surveillance radars in the northeastern USA.
Ecological Applications 26(3): 752-770.
Feller K, Lagerholm S, Clubwala R, et al. (2009) Characterization of photoreceptor cell types
in the little brown bat Myotis lucifugus (Vespertilionidae), Comparative Biochemistry and
Physiology Part B, Biochemistry & Molecular Biology 154: 412418.
Fischer JH, Debski I, Taylor GA, et al. (2021) Consistent offshore artificial light at night near
the last breeding colony of a critically endangered seabird. Conservation Science and
Practice 3(9): e481.
Fobert EK, Burke da Silva K and Swearer SE (2019) Artificial light at night causes
reproductive failure in clownfish. Biology Letters 15:e20190272.
Foster, RG and Kreitzman, L (2005) Rhythms of Life. Profile Books, London.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
129
Frick, WF, Kingston, T and Flanders J (2020) A review of the major threats and challenges
to global bat conservation. Annals of the New York Academy of Sciences 1469: 5-25.
Fritches KA (2012) Australian loggerhead sea turtle hatchlings do not avoid yellow. Marine
and Freshwater Behaviour and Physiology 45(2):79-89.
Gastman EA (1886) Birds killed by electric light towers at Decatur, Ill. American
Naturalist, 20(11), p.981.
Gaston KJ, Visser ME and Holker F (2018) The biological impacts of artificial light at night:
the research challenge. Philosophical Transactions of the Royal Society B 370:e20140133.
Gauthreaux SA and Belser CG (2006) Effects of artificial night lighting on migrating birds. In:
Ecological Consequences of Artificial Night Lighting, Rich C and Longcore T, Editors. Island
Press: Washington, D.C., USA, p:67-93.
Gehring J, Kerlinger P and Manville II AM (2009) Communication towers, lights, and birds:
successful methods of reducing the frequency of avian collisions. Ecological Applications
19(2): 505514.
Gillings S and Scott C (2021) Nocturnal flight calling behaviour of thrushes in relation to
artificial light at night. Ibis163(4): 1379-1393.
Gineste B, Souquet M, Couzi F-X, et al. (2016) Tropical shearwater population stability at
Reunion Island, despite light pollution. Journal of Ornithology 158:385-394.
González-Bernal E, Brown G and Shine R (2014) Invasive cane toads: Social facilitation
. PLoS ONE 9(7):e102880.
Griesemer AM and Holmes ND (2011) Newell’s shearwater population modeling for Habitat
Conservation Plan and Recovery Planning Technical Report No. 176. The Hawai`i- Pacific
Islands Cooperative Ecosystem Studies Unit and Pacific Cooperative Studies Unit University
of Hawai`i: Honolulu, Hawai`i. 68.
Grubisic M, Haim A, Bhusal P, et al. (2019) Light Pollution, Circadian Photoreception, and
Melatonin in Vertebrates. Sustainability 11: 6400.
Günther A, Einwich A, Sjulstok E, et al. (2018) Double-cone localization and seasonal
expression pattern suggest a role in magnetoreception for European robin Cryptochrome 4.
Current Biology 28(2): P211-223.
Haddock JK (2018) Effects of artificial lighting on insectivorous bat communities in urban
ecosystems. A thesis submitted in fulfilment of the requirements for the degree of Doctor of
Philosophy in the School of Life and Environmental Sciences, Faculty of Science, at The
University of Sydney.
Haddock JK, Threlfall CG, Law B, et al. (2019a) Light pollution at the urban forest edge
negatively insectivorous bats. Biological Conservation 236: 17-28.
Haddock JK, Threlfall CG, Law B, et al. (2019b) Responses of insectivorous bats and
nocturnal insects to local changes in street light technology. Austral Ecology 44(6): 1052-
1064.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
130
Hnel A, Posch T, Ribas SJ, et al. (2018) Measuring night sky brightness: methods and
challenges. Journal of Quantitative Spectroscopy and Radiative Transfer doi:
10.1016/j.jqsrt.2017.09.008
Harewood A and Horrocks J (2008) Impacts of coastal development on hawksbill hatchling
survival and swimming success during the initial offshore migration. Biological Conservation
141:394-401.
Hart NS (2001) The visual ecology of avian photoreceptors. Progress in Retinal and Eye
Research 20:675-703.
Harvie-Brown JA (1880) Report on the Migration of Birds. West, Newman.
Henderson PA and Southwood TRE (2016) Ecological Methods 4th Edition.: Wiley-
Blackwell. p. 656.
Hodge W, Limpus CJ and Smissen P (2007) Queensland turtle conservation project:
Hummock Hill Island Nesting Turtle Study December 2006 Conservation Technical and Data
Report Environmental Protection Agency, Queensland. 1-10.
Hooker D (1911) Certain reactions to color in the young loggerhead turtle. Papers from the
Tortugas Laboratory - Carnegie Institute 13:71-76.
Horch KW, Gocke JP, Salmon M, et al. (2008) Visual spectral sensitivity of hatchling
loggerhead (Caretta caretta L.) and leatherback (Dermochelys coriacea L.) sea turtles, as
determined by single-flash electroretinography. Marine and Freshwater Behaviour and
Physiology 41(2):107-119.
Horton KG, Van Doren BM, Stepanian PM et al. (2016a) Seasonal differences in landbird
migration strategies. Auk 133: 761769
Horton KG, Van Doren BM, Stepanian PM et al. (2016b) Where in the air? Aerial habitat use
of nocturnally migrating birds. Biology Letters 12: 20160591
Horton KG, Nilsson C, Van Doren BM, et al. (2019a) Bright lights in the big cities: migratory
Frontiers in Ecology and the Environment 17(4): 209-214.
Horton KG, Van Doren BM, La Sorte FA, et al. (2019b) Holding steady: Little change in
intensity or timing of bird migration over the Gulf of Mexico. Global Change Biology 25(3):
1106-1118.
Horton KG, Van Doren BM, Albers HJ, et al. (2021) Near-term ecological forecasting for
dynamic aeroconservation of migratory birds. Conservation Biology 35(6): 1777-1786.
Hoyos-Díaz JM, Villalba-Alemán E, Ramoni-Perazzi P, et al. (2018). Impact of artificial
lighting on capture success in two species of frugivorous bats (Chiroptera: Phyllostomidae)
in an urban locality from the Venezuelan Andes. Mastozoologia Neotropical 25(2): 473-478.
Hu Z, Hu H and Huang Y (2018) Association between nighttime artificial light pollution and
sea turtle nest density along Florida coast: A geospatial study using VIIRS remote sensing
data. Environmental Pollution 239:30-42.
IDA (International Dark-Sky Association) (2019) Guidance for Electronic Message Centers
(EMCs). Available at: https://www.darksky.org/wp-content/uploads/2019/05/EMC-Guidelines-
IDA2019-1.pdf

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
131
IDA (International Dark-Sky Association) and IES (Illuminating Engineering Society) (2020)
Five Principles for Responsible Outdoor Lighting. Available at: https://www.darksky.org/our-
work/lighting/lighting-principles/
Imamoto Y and Shichida Y (2014) Cone visual pigments. Biochim. Biophys. Acta 1837:664
673.
Imber MJ (1975) Behaviour of petrels in relation to the moon and artificial lights. Notornis
22:302-306.
Irsitech (2018) https://iristech.co/how-iris-reduces-blue-light/visible-spectrum. 2018 [cited
Accessed 1st October 2018.
ISO/CIE 23539:2023 Photometry The CIE System of Physical Photometry. Available at:
https://cie.co.at/publications/photometry-cie-system-physical-photometry-3
IUCN (2023) The IUCN Red List of Threatened Species. Available at:
https://www.iucnredlist.org
Jechow A, Ribas SJ, Domingo RC, et al. (2018) Tracking the dynamics of skyglow with
differential photometry using a digital camera with fisheye lens. Journal of Quantitative
Spectroscopy and Radiative Transfer 209:212-223.
Jechow A, Kyba CCM and Hlker F (2019) Beyond All-Sky: Assessing Ecological Light
Pollution Using Multi-Spectral Full-Sphere Fisheye Lens Imaging Journal of Imaging
5(46):doi:10.3390/jimaging5040046.
Jones J and Francis CM (2003) The effects of light characteristics on avian mortality at
lighthouses. Journal of Avian Biology 34: 328-333.
Kamrowski RL, Limpus CJ, Moloney J, et al. (2012) Coastal light pollution and marine
turtles: Assessing the magnitude of the problem. Endangered Species Research 19:85-98.
Kamrowski RL, CJ L, Pendoley K and Hamann M (2014) Influence of industrial light pollution
on the sea-finding behaviour of flatback turtle hatchlings. Wildlife Research 41:421-434.
Kerbiriou C, Barré K, Mariton L, et al. (2020) Switching LPS to LED streetlight may
dramatically reduce activity and foraging of bats. Diversity 12(4): 165.
Kemp MU, Shamoun-Baranes J, Dokter AM, et al. (2013) The influence of weather on the
flight altitude of nocturnal migrants in mid-latitudes. Ibis 155: 734749.
Kernbach ME, Cassone VM, Unnasch TR, et al. (2020) Broad-spectrum light pollution
suppresses melatonin and increases West Nile virusinduced mortality in House Sparrows
(Passer domesticus). The Condor 122(3): duaa018.
Kirby JS, Stattersfield AJ, Butchart SHM, et al. (2008) Key conservation issues for migratory
land- and waterbird species on the worlBird Conservation International 18:
S49-S73.
Klem Jr D (2008) Avian mortality at windows: the second largest human source of bird
mortality on Earth. Proceedings of the Fourth International Partners in Flight Conference:
Tundra to Tropics. 244-251

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
132
Kollth Z (2010) Measuring and modelling light pollution at the Zselic Starry Sky Park.
Journal of Physics: Conference Series 2018(5th Workshop of Young Researchers in
Astronomy and Astrophysics):012001.
study of light pollution. Journal of Ornithology 163: 827-841.
Kreitzman L and Foster RG (2010) Seasons of Life. Profile Books, London.
Kruskop SV (2021) Diversity Aspects in Bats: Genetics, Morphology, Community Structure.
Diversity 13: 424.
Kumar R, Prasad DN and Elangovan V (2018) The effect of seasonal changes on
emergence behaviour of the Indian flying fox, Pteropus giganteus. Proceedings of the
Zoological Society. 72: 74-78.
Kyba CCM, Ruhtz T, Fishcher J, et al. (2011) Cloud coverage acts as an amplifier for
ecological light pollution in urban ecosystems. PLoS ONE 6(e17307).
Kyba CCM, Kuester T, Snchez de Miguel A, et al. (2017) Artificially lit surface of Earth at
night increasing in radiance and extent. Science Advances 3:e1701528.
Kyba CCM, Mohar A, Pintar G, et al. (2018) Reducing the environmental footprint of church
lighting: matching façade shape and lowering luminance with the EcoSky LED. International
Journal of Sustainable Lighting 20(1): 1-10.
Kyheröinen E-M, Aulagnier S, Dekker J, et al. (2019) Guidance on the conservation and
management of critical feeding areas and commuting routes for bats. EUROBATS
Publication Series No. 9. UNEP/EUROBATS Secretariat, Bonn, Germany, 109 pp. Available
at:
https://www.eurobats.org/sites/default/files/documents/publications/publication_series/WEB_
DIN_A4_EUROBATS_09_ENGL_NVK_01042019.pdf
La Sorte FA, Fink D, Hochachka WM, et al. (2014) The role of atmospheric conditions in the
seasonal dynamics of North American migration flyways. Journal of Biogeography 41: 1685-
1696.
La Sorte FA, Hochachka WM, Farnsworth A, et al. (2015) Migration timing and its
determinants for nocturnal migratory birds during autumn migration. Journal of Animal
Ecology 84: 12021212.
La Sorte FA, Fink D, Buler JJ, et al. (2017) Seasonal associations with urban light pollution
for nocturnally migrating bird populations. Global Change Biology, 23(11), pp.4609-4619.
La Sorte FA and Horton KG (2021) Seasonal variation in the effects of artificial light at night
on the occurrence of nocturnally migrating birds in urban areas. Environmental
Pollution, 270, p.116085.
La Sorte FA, Aronson MFJ, Lepczyk CA, et al. (2022) Assessing the combined threats of
Global
Ecology and Biogeography In press
Lacoeuilhe A, Machon N, Julien J-F, et al. (2014) The Influence of Low Intensities of Light
Pollution on Bat Communities in a Semi-Natural Context. PLoS ONE. 9(10): e103042.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
133
Lao S, Robertson BA, Anderson AW, et al. (2020) The influence of artificial light at night and
polarized light on bird-building collisions. Biological Conservation 241: 108358.
Lao S, Anderson AW, Blair RB, et al. (2023) Birdbuilding collisions increase with weather
conditions that favor nocturnal migration and with inclement and changing
weather. Ornithological Applications, 125(1), p.duac045.
Le Corre M, Ollivier A, Ribes S et al. (2002) Light-induced mortality of petrels: a 4-year study
from Runion Island (Indian Ocean). Biological Conservation 105:93-102.
Lebbin DJ, Harvey MG, Lenz TC, et al. (2007) Nocturnal Migrants Foraging at Night by
Artificial Light. The Wilson Journal of Ornithology 119(3): 506-508.
Lee KEM, Lum WHD and Coleman JL (2021) Ecological impacts of the LED-streetlight
retrofit on insectivorous bats in Singapore. PLoS ONE 16(5): e0247900.
Levenson DH, Eckert SA, Crognale MA, et al. (2004) Photopic Spectral Sensitivity of Green
and Loggerhead Sea Turtles. Copeia 2004(2):908-914.
Levin N, Kyba CCM, Zhang Q, et al. (2020) Remote sensing of night lights: A review and an
outlook for the future. Remote Sensing of the Environment 237:111443.
Lewanzik D and Voigt CC (2014) Artificial light puts ecosystem services of frugivorous bats
at risk. Journal of Applied Ecology 51(2): 388-394.
Limpus CJ, Miller JD, Parmenter CJ, et al. (2003) The green turtle, Chelonia mydas,
population of Raine Island and the Northern Great Barrier Reef: 1843-2001. Memoirs of the
Queensland Museum 49:349-440.
Limpus CJ and Kamrowski RL (2013) Ocean-finding in marine turtles: The importance of low
horizon elevation as an orientation cue. Behaviour 150:863-893.
Lohmann KJ, Witherington B, Lohmann CMF, et al. (1997) Orientation, navigation, and natal
beach homing in sea turtles. In: The Biology of Sea Turtles. Volume I, Lutz PL and Musick
JA, Editors. CRC Press: Washington D.C., p:107-135.
Longcore T and Rich C (2004) Ecological light pollution. Frontiers in Ecology and the
Environment, 2(4), pp.191-198.
Longcore T, Rich C, Mineau P, et al. (2013) Avian mortality at communication towers in the
United States and Canada: which species, how many, and where? Biological Conservation
158:410-419.
Longcore T, Aldern HL, Eggers JF, et al. (2015) Tuning the white light spectrum of light
emitting diode lamps to reduce attraction of nocturnal arthropods. Philosophical Transactions
Royal Society B 370: 201401125.
Longcore T, Rodrguez A, Witherington B, et al. (2018) Rapid assessment of lamp spectrum
to quantify ecological effects of light at night. Journal of Experimental Zoology Part A
Ecological and Integrative Physiology 2018:1-11.
Loss SR, Will T, Loss SS, et al. (2014) Bird-building collisions in the United States:
Estimates of annual mortality and species vulnerability. The Condor 116: 8-23.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
134
Loss SR, Lao S, Eckles JW, et al. (2019) Factors influencing bird-building collisions in the
downtown area of a major North American city. PLoS ONE 14(11): e0224164.
Loss SR, Li BV, Horn LC, et al (2023) Citizen Science to address the global issue of bird-
window collisions. Frontiers in Ecology and the Environment https://doi.org/10.1002/fee.2614
Loureno PM, Silva A, Santos CD, et al. (2008) The energetic importance of night foraging
for waders wintering in a temperate estuary. Acta Oecologica 34:122-139.
Luo B, Xu R, Li Y, et al. (2021) Artificial light reduces foraging opportunities in wild least
horseshoe bats. Environmental Pollution 288: 117765.
McLaren JD, Buler JJ, Schreckengost T, et al. (2018) Artificial light at night confounds broad-
scale habitat use by migrating birds. Ecology Letters 21(3):356-364.
McNeil R, Drapeau P and Pierotti R (1993) Nocturnality in Colonial Waterbirds: Occurrence,
Special Adaptations, and Suspected Benefits'. In: Current Ornithology, Power DM, Editor.
Springer US: Boston, MA, p:187-246.
Mena JL, Rivero J, Bonifaz E, et al. (2021) The effect of artificial light on bat richness and
nocturnal soundscapes along an urbanization gradient in an arid landscape of central Peru.
Urban Ecosystems. doi: 10.1007/s11252-021-01163-x
Merkel FR and Johansen KL (2011) Light-induced bird strikes on vessels in Southwest
Greenland. Marine Pollution Bulletin 62:2330-2336.
Mitkus M, Nevitt GA, Danielsen J, et al. (2016) Vision on the high seas: spatial resolution
and optical sensitivity in two procellariform seabirds with different foraging strategies. Journal
of Experimental Biology 219:3329-3338.
Montevecchi WA (2006) Influences of Artificial Light on Marine Birds. In: Ecological
consequences of artificial night lighting, Rich C and Longcore T, Editors. Island Press:
Washington DC, p:480.
Moreau RE (1972) The Palaearctic-African bird migration systems. London, New York,
Academic Press.
Moro D, van de Merwe J, Thomas M, et al. (2018) Integrating resource development with
island conservation: Barrow Island as a model for conservation and development. In:
Australian Island Arks: Conservation, Management and Opportunities, Moro D, Ball D and
Bryant S, Editors. CSIRO Publishing: Melbourne, p:131-146.
Mrosovsky N (1968) Nocturnal emergence of hatchling sea turtles: control by thermal
inhibition of activity. Nature 220:1338-1339.
Mrosovsky N (1972) The water finding ability of sea turtles. Brain Behaviour and Evolution
5:202-225.
Mrosovsky N and Shettleworth SJ (1968) Wavelength preferences and brightness cues in
the water finding behaviour of sea turtles. Behaviour 32:211-257.
Mu T, Cai S, Peng H-B, et al. (2022) Evaluating staging habitat quality to advance the
conservation of a declining migratory shorebird, Red Knot Calidris canutus. Journal of
Applied Ecology 59(8): 2084-2093.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
135
Murugavel B, Kelber A and Somanathan H (2021) Light, flight and the night: effect of
ambient light and moon phase on flight activity of pteropodid bats. Journal of Comparative
Physiology 207: 59-68.
Murphy RC (1936) Oceanic birds of South America. New York: Macmillan. p.
Newman EA and Hartline PH (1981) Integration of visual and infrared information to
biomodal neurons in the rattlesnake optic tectum. Science 213(4509):789-91.
Newton I (2007) Weather-related mass-mortality events in migrants. Ibis 149: 453-467.
Newton I (2008) The migration ecology of birds. Academic Press, London.
Nichols KS, Homayoun T, Eckles J, et al. (2018) Bird-building collision risk: An assessment
of the collision risk of birds with buildings by phylogeny and behavior using two citizen-
science datasets. PLoS ONE 13(8): e0201558.
Norevik G, Åkesson S, Andersson A, et al. (2019) The lunar cycle drives migration of a
nocturnal bird. PLoS Biology 17(10): e3000456.
Ouyang JQ, de Jong M, Hau M, et al. (2015) Stressful colours: corticosterone concentrations
in a free-living songbird vary with the spectral composition of experimental illumination.
Biology Letters 11:20150517.
Owens ACS and Lewis SM (2018) The impact of artificial light at night on nocturnal insects:
A review and synthesis. Ecology and Evolution 8(22): 11337-11358.
Owens ACS, Cochard P, Durrant J, et al. (2020) Light pollution is a driver of insect declines.
Biological Conservation 241: 108259.
Parkins KL, Elbin SB and Barnes E (2015) Light, glass, and bird-building collisions in an
urban park. Northeastern Naturalist 22(1): 84-94.
Pendoley K (2000) The influence of gas flares on the orientation of Green Turtle hatchlings
at Thevenard Island, Western Australia in Pilcher NJ and Ismail G, Editors, Second ASEAN
Symposium and Workshop on Sea Turtle biology and Conservation ASEAN Academic
Press: Kota Kinabalu, Borneo. 130-142.
Pendoley KL (2005) Sea Turtles and the Environmental Management of Industrial Activities
in North Western Australia Murdoch University. 330.
Pendoley K and Kamrowski RL (2015a) Influence of horizon elevation on the sea-finding
behaviour of hatchling flatback turtles exposed to artificial light glow. Marine Ecology
Progress Series 529:279-288.
Pendoley K and Kamrowski RL (2015b) Sea-finding in marine turtle hatchlings: What is an
appropriate exclusion zone to limit disruptive impacts of industrial light at night? Journal for
Nature Conservation 30:1-11.
Pendoley KL, Whittock PA, Vitenbergs A, et al. (2016) Twenty years of turtle tracks: marine
turtle nesting activity at remote locations in the Pilbara, Western Australia. Australian Journal
of Zoology 64:217-226.
Pennell JP (2000) The Effect of Filtered Roadway Lighting on Nesting by Loggerehad Sea
Turtles (Caretta caretta) and Green Turtle (Chelonia mydas) Hatchlings Florida Atlantic
University: Boca Raton.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
136
Piersma T and Baker AJ (2000) Life history characteristics and the conservation of migratory
shorebirds. In: Behaviour and conservation, Gosling LM and Sutherland WJ, Editors.
Cambridge University Press: Cambridge, United Kingdom, p:105-124.
Piersma T, Lok T, Chen Y, et al. (2016) Simultaneous declines in summer survival of three
shorebird species signals a flyway at risk. Journal of Applied Ecology 53(2): 479-490.
Pinzon-Rodriguez A, Bensch S and Muheim R (2018) Expression patterns of cryptochrome
genes in avian retina suggest involvement of Cry4 in light-dependent magnetoreception.
Journal of the Royal Society Interface 15: 20180058.
collisions with urban structures on Kauai. Colonial Waterbirds 21:20-34.
Poot H, Ens B, Vries H, et al. (2008) Green light for nocturnally migrating birds. Ecology and
Society 13(2):47.
Price JT, Drye B, Domangue RJ, et al. (2018) Exploring the role of artificial light in
Loggerhead turtle (Caretta caretta) nest-site selection and hatchling disorientation.
Herpetological Conservation and Biology 13(2):415-422.
S
distributions. Journal of Biogeography 32: 607-614.
Raine H, Borg JJ, Raine A, et al. (2007) Light Pollution and Its Effect on Yelkouan
Shearwaters in Malta; Causes and Solutions BirdLife Malta: Malta: Life Project Yelkouan
Shearwater. 1-54.
Rebke M, Dierschke V, Weiner CN, et al. (2019) Attraction of nocturnally migrating birds to
artificial light: The influence of colour, intensity and blinking mode under different cloud cover
conditions. Biological Conservation 233: 220-227.
Reed JR, Sincock JL and Hailman JP (1985) Light attraction in endangered procellariform
birds: reduction by shielding upward radiation. Auk 102:377-383.
Reed JR (1986) Seabird vision: Spectral sensitivity and light-attraction behavior University of
Wisconsin: Madison, Wisconsin. 190.
Rich C and Longcore T (eds.) (2006) Ecological consequences of artificial night lighting.
Island Press: Washington DC. 480.
Richardson WJ (1978) Timing and amount of bird migration in relation to weather: A
review. Oikos 30: 224272.
-scale temporal variation in bird-window
collisions in the central United States. Scientific Reports, 11(1), p.11062.
Robert KA, Lesku JA, Partecke J, et al. (2015) Artificial light at night desynchronizes strictly
seasonal reproduction in a wild mammal. Proceedings of the Royal Society B 282:20151745.
-f
sea-finding behaviour of loggerhead turtle hatchlings (Caretta caretta). Wildlife Research
43:27-37.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
137
Rodrguez A, Rodrguez B and Lucas MP (2012) Trends in numbers of petrels attracted to
artificial lights suggest population declines in Tenerife, Canary Islands. Ibis 154:167-172.
Rodrguez A, Burgan G, Dann P, et al. (2014) Fatal attraction of short-tailed shearwaters to
artificial lights. PLoS ONE 9(10):e110114.
a D, Rodrguez B, et al. (2015a) Artificial lights and seabirds: is light
pollution a threat for the threatened Balearic petrels? Journal of Ornithology 156:893-902.
Rodrguez A, Rodrguez B and Negro JJ (2015b) GPS tracking for mapping seabird mortality
induced by light pollution. Scientific Reports 5:10670.
Rodrguez A, Dann P and Chiaradia A (2017a) Reducing light-induced mortality of seabirds:
High pressure sodium lights decrease the fatal attraction of shearwaters. Journal for Nature
Conservation 39:68-72.
Rodríguez A, Moffet J, Revoltos A, et al. (2017b) Light pollution and seabird fledglings:
targeting efforts in rescue programs. Journal of Wildlife Management 81:734-741.
Rodrguez A, Holmes ND, Ryan PG, et al. (2017c) A global review of seabird mortality
caused by land-based artificial lights. Conservation Biology 31:986-1001.
Rodrguez A, Holmberg R, Dann P, et al. (2018) Penguin colony attendance under artificial
lights for ecotourism. JEZ-A Ecological and Intergrative Physiology 329(8- 9):457-464.
Rodríguez A, Rodríguez B, Acosta Y, et al. (2022) Tracking flights to investigate seabird
mortality induced by artificial lights. Frontiers in Ecology and Evolution 9: 786557.
Rogers DI, Battley PF, Piersma T, et al. (2006a) High-tide habitat choice: insights from
modelling roost selection by shorebirds around a tropical bay. Animal Behaviour 72(3):563-
575.
Rogers DI, Piersma T and Hassell CJ (2006b) Roost availability may constrain shorebird
distribution: Exploring the energetic costs of roosting and disturbance around a tropical bay.
Biological Conservation 133(2):225-235.
Rojas LM, McNeil R, Cabana T, et al. (1999) Diurnal and nocturnal visual capabilities in
shorebirds as a function of their feeding strategies. Brain Behavior and Evolution 53(1):29-
43.
Ross GJB, Burbidge AA, Canty P, et al. (1996) Status of Australia's Seabirds. In: State of
the Environment Report. CSIRO Sustainable Ecosystems: Perth, p:167-182.
Rowse EG, Lewanzik D, Stone EL, et al. (2016) Dark matters: the effects of artificial lighting
on bats. In C.C Voigt and T. Kingston (eds.) Bats in the anthropocene: conservation of bats
in a changing world. pp. 187213. New York, NY: Springer (Open access;
http://www.springer.com/gb/book/ 9783319252186).
Russart KLG and Nelson RJ (2018) Artificial light at night alters behavior in laboratory and
wild animals. JEZ-A Ecological and Integrative Physiology 329(8-9):401-408.
Russo D, Cistrone L, Libralato N, et al. (2017). Adverse effects of artificial illumination on bat
drinking activity. Animal Conservation 20(6): 492-501.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
138
Russo D, Cosentino F, Festa F, et al. (2019). Artificial illumination near rivers may alter bat-
insect trophic interactions. Environmental Pollution 252: 1671-1677.
Rydell J (1992) Exploitation of insects around streetlamps by bats in Sweden. Functional
Ecology 6(6): 744-750.
Rydell J, Eklöf J and Sánchez-Navarro S (2017) Age of enlightenment: long-term effects of
outdoor aesthetic lights on bats in churches. Royal Society Open Science 4: 161077.
Rydell J, Michaelsen TC, Sánchez-Navarro S, et al. (2021) How to leave the church: light
avoidance by brown long-eared bats. Mammalian Biology 101: 979-986.
Salinas-Ramos VB, Ancillotto L, Cistrone L, et al. (2021) Artificial illumination influences
niche segregation in bats. Environmental Pollution 284: 117187.
Salmon M (2003) Artificial night lighting and sea turtles. Biologist 50:163-168.
Salmon M (2006) Protecting Sea Turtles from Artificial Night Lighting at Florida's Oceanic
Beaches. In: Ecological Consequences of Artificial Night Lighting, Rich C and Longcore
T,(eds.) Island Press: Washinton DC, p:141-168.
Salmon M and Wyneken J (1990) Do swimming loggerhead turtles (Caretta caretta L.) use
light cues for offshore orientation? . Marine Behavioural Physiology 17:233-246.
Salmon M, Wyneken J, Fritz E, et al. (1992) Sea finding by hatchling sea turtles: Role of
brightness, silhouette and beach slope as orientation cues. Behaviour 122:56- 77.
Salmon M, Reiners R, Lavin C, et al. (1995) Behavior of loggerhead sea turtles on an urban
beach. I. Correlates of nest placement. Journal of Herpetology 29(4):560- 567.
Sánchez de Miguel A, Bennie J, Rosenfeld E, et al. (2021) First Estimation of Global Trends
in Nocturnal Power Emissions Reveals Acceleration of Light Pollution. Remote Sensing
13(16): 3311.
Sanders D and Gaston KJ (2018) How ecological communities respond to artificial light at
night. Journal of Experimental Zoology 329(8-9):394-400.
Sanders D, Frago E, Kehoe R, et al. (2021) A meta-analysis of biological impacts of artificial
light at night. Nature Ecology and Evolution 5: 74-81.
Santiago-Quesada F, Estrella SM, Sanchez-Guzman JM et al. (2014) Why water birds
forage at night: a test using black-tailed godwits Limosa limosa during migratory periods.
Journal of Avian Biology 45(4):406-409.
Santos CD, Miranda AC, Granadeiro JP, et al. (2010) Effects of artificial illumination on the
nocturnal foraging of waders. Acta Oecologica 36:166-172.
Schneider RM, Barton CM, Zirkle KW, et al. (2018) Year-round monitoring reveals
prevalence of fatal bird-window collisions at the Virginia Tech Corporate Research Center.
PeerJ 6: e4562.
Scott KM, Danko A, Plant P, et al. (2023) What causes bird‐building collision risk? Seasonal
dynamics and weather drivers. Ecology and Evolution, 13(4), p.e9974.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
139
Seewagen CL and Adams AM (2021) Turning to the dark side: LED light at night alters the
activity and species composition of a foraging bat assemblage in the northeastern United
States. Ecology and Evolution 11: 5635-5645.
Senzaki M, Barber JR, Phillips JN, et al. (2020) Sensory pollutants alter bird phenology and
fitness across a continent. Nature 587 (7835):605-609.
Shcherbakov S, Knörzer A, Espenhahn S, et al. (2013) Sensitivity Differences in Fish Offer
Near-Infrared Vision as an Adaptable Evolutionary Trait. PLoS ONE 8(5): e64429.
Simmonds NB and Cirranello AL (2023) Bat species of the World: A taxonomic and
geographic database. Version 1.3. Available at: https://www.batnames.org
Simões BF, Foley NM, Hughes GM, et al. (2018) As blind as a bat? Opsin phylogenetics
illuminates the evolution of color vision in bats. Molecular Biology and Evolution 36: 5468.
Smith RA, Gagné M and Fraser KC (2021) Pre-migration artificial light at night advances the
spring migration timing of a trans-hemispheric migratory songbird. Environmental Pollution
269: 116136.
Somers-Yeates R, Hodgson D, McGregor PK, et al. (2013) Shedding light on moths: shorter
wavelengths attract noctuids more than geometrids. Biology Letters 9: 20130376.
Spoelstra K and Visser ME (2013) The impact of artificial light on avian ecology. Avian
Urban Ecol, 4, pp.21-28.
Spoelstra K, van Grunsven RHA, Ramakers JJC, et al. (2017) Response of bats to light with
different spectra: light-shy and agile bat presence is affected by white and green, but not red
light. Proc. R. Soc. B. 284: 20170075.
Spoelstra K, Ramakers JJC, van Dis NE, et al. (2018) No effect of artificial light of different
Myotis daubentonii) in a choice experiment. Journal
of Experimental Zoology Part A: Ecological Genetics and Physiology 329: 506-510.
State of Queensland (2020) Flying-fox roost management guideline. Prepared by: Wildlife
and Threatened Species Operations, Department of Environment and Science. Available at:
https://www.qld.gov.au/__data/assets/pdf_file/0009/221022/Guideline-Roost-
Management.pdf
Stone EL, Harris S and Jones G (2015) Impacts of artificial lighting on bats: a review of
challenges and solutions. Mammalian Biology 80: 213-219.
Straka TM, Wolf M, Gras P, et al. (2019) Tree cover mediates the effect of artificial light on
urban bats. Frontiers in Ecology and Evolution 7: 91.
Straka TM, Greif S, Schultz S, et al. (2020) The effect of cave illumination on bats. Global
Ecology and Conservation 21: e00808.
Straka TM, von der Lippe M, Voigt CC, et al. (2021) Light pollution impairs urban nocturnal
pollinators but less so in areas with high tree cover. Science of the Total Environment 778:
146244.
Surman CA and Nicholson LW (2014a) The Integrated Shearwater Monitoring Project
(ISMP): Annual Report for the 2013/14 Season. Unpublished report prepared for Apache
Energy Ltd. Halfmoon Biosciences. 47.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
140
Surman CA and Nicholson LW (2014b) Monitoring of annual variation in seabird breeding
colonies throughout the Lowendal Group of islands: 2014 Annual Report. Lowendal Island
Seabird Monitoring Program (LISMP) Unpublished report prepared for Apache Energy Ltd.
by Halfmoon Biosciences. 59.
Syposz M, Goncalves F, Carty M, et al. (2018) Factors influencing Manx Shearwater
grounding on the west coast of Scotland. Ibis 160:846-854.
Syposz M, Padget O, Willis J, et al. (2021) Avoidance of different durations, colours and
intensities of artificial light by adult seabirds. Scientific Reports 11: 18941.
Szaz D, Horvath G, Barta A, et al. (2015) Lamp-lit Bridges as Dual Light-Traps for the Night-
Swarming Mayfly, Ephoron virgo: Interaction of Polarized and Unpolarized Light Pollution.
PLoS ONE 10(3): e0121194.
Tanalgo KC, Tabora JAG, Oliveira HFM, et al. (2021) DarkCides 1.0, a global database for
bats in karsts and caves. Scientific Data 9: 155.
Telfer TC, Sincock JL, Byrd GV, et al. (1987) Attraction of Hawaiian seabirds to lights:
conservation efforts and effects of moon phase. Wildlife Society Bulletin 15:406- 413.
Thums M, Whiting SD, Reisser JW, et al. (2016) Artificial light on water attracts turtle
hatchlings during their near shore transit. Royal Society Open Science 3:e160142.
Tosini G, Ferguson I and Tsubota K (2016) Effects of blue light on the circadian system and
eye physiology. Molecular Vision 22:61-72.
Troy J, Holmes N, Veech J, et al. (2013) Using observed seabird fallout records to infer
patterns of attraction to artificial light. Endangered Species Research 22:225-234.
Ulrich W, Sachanowicz K and Michalak M (2007) Environmental correlates of species
richness of European bats (Mammalia: Chiroptera) Acta Chiropterologica 9(2): 347-360.
(UN) United Nations (2019) World Urbanization Prospects: The 2018 Revision
(ST/ESA/SER.A/420) Department of Economic and Social Affairs, Population Division. New
York: United Nations. Available at:
https://population.un.org/wup/Publications/Files/WUP2018-Report.pdf
UNEP/AEWA Secretariat (2021) Status and trends of migratory waterbird populations in the
African-Eurasian Flyways. AEWA Popular Series. Bonn, Germany. Available at:
https://www.unep-
aewa.org/sites/default/files/publication/AEWA%20Conservation%20Status%20Report_FINA
L_EN_WEB.pdf
UNEP/CMS (2014) A Review of Migratory Bird Flyways and Priorities for Management.
UNEP/CMS Secretariat, Bonn, Germany, 164 pages. CMS Technical Series No. 27.
Available at:
https://www.cms.int/sites/default/files/publication/CMS_Flyways_Reviews_Web.pdf
UNOOSA (United Nations Office for Outer Space Affairs) (2020) Dark and Quiet Skies for
Science and Society. On-line Workshop Report and Recommendations. Available at:
https://www.iau.org/static/publications/dqskies-book-29-12-20.pdf
UNOOSA (United Nations Office for Outer Space Affairs) (2021) Dark and Quiet Skies for
Science and Society II. On-line Workshop Report and Recommendations. Available at:
https://www.iau.org/static/science/scientific_bodies/working_groups/286/dark-quiet-skies-2-
working-groups-reports.pdf

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
141
Van Belle J, Shamoun-Baranes J, Van Loon E, et al. (2007) An operational model predicting
autumn bird migration intensities for flight safety. Journal of Applied Ecology 44: 864874.
Van de Kam J, Ens B, Piersma T, et al. (2004) Shorebirds: an illustrated behavioural
ecology. Utrecht, the Netherlands: KNNV Publishers. p. 368.
Van Doren BM, Sheldon D, Geevarghese J, et al. (2015) Autumn morning flights of migrant
songbirds in the northeastern United States are linked to nocturnal migration and winds
aloft. Auk 132:105118
Van Doren BM, Horton KG, Dokter AM, et al. (2017) High-intensity urban light installation
dramatically alters nocturnal bird migration. Proceedings of the National Academy of
Sciences, 114(42), pp.11175-11180.
Van Doren BM and Horton KG (2018) A continental system for forecasting bird migration.
Science 361(6407): 1115-1118.
Van Doren BM, Willard DE, Hennen M, et at. (2021) Drivers of fatal bird collisions in an
urban center. PNAS 118(24): e2101666118.
Verheijen FJ (1985) Photopollution - artificial light optic spatial control systems fail to cope
with incidents, causations, remidies. Experimental Biology 44(1):1-18.
Voigt CC and Kingston T (eds.) (2016) Bats in the Anthropocene: Conservation of Bats in a
Changing World. Springer Cham.
Voigt CC, Azam C, Dekker J et al. (2018a) Guidelines for consideration of bats in lighting
projects. EUROBATS Publication Series No. 8 UNEP/EUROBATS Secretariat, Bonn,
Germany, 62.pp.
Voigt CC, Rehnig K, Lindecke O et al. (2018b) Migratory bats are attracted by red light but
not by warm-white light: Implications for the protection of nocturnal migrants. Ecology and
Evolution 8: 9353-9361. doi: 10.1002/ece3.4400
Voigt CC, Dekker J, Fritze M, et al. (2021) The impact of light pollution on bats varies
according to foraging guild and habitat context. BioScience 71(10): 1103-1109.
Vorobyev M (2003) Coloured oil droplets enhance colour discrimination. Proceedings
Biological Sciences 270:12551261.
Votier SC and Sherley RB (2017) Seabirds. Current Biology 27: R448-R450.
https://www.cell.com/current-biology/pdf/S0960-9822(17)30075-1.pdf
Wakefield A, Broyles M, Stone EL, et al. (2018) Quantifying the attractiveness of broad-
spectrum street lights to aerial nocturnal insects. Journal of Applied Ecology 55: 714-722.
doi: 10.1111/1365-2664.13004
Warham J (1990) The Behaviour, Population Biology and Physiology of the Petrels. London:
Academic Press. p. 440.
Warrant E (2019) Invertebrates vision. Encyclopedia of Animal Behaviour, 2nd Edn, ed. J. C.
Choe (Amsterdam: Elsevier), 6479.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
142
Warrant EJ, Frost B, Green K, et al. (2016) The Australian Bogong moth Agrotis infusa: A
long-distance nocturnal navigator. Frontiers in Behavioural Neuroscience 10:doi:
10.3389/fnbeh.2016.00077.
Watanuki Y (1986) Moonlight avoidance behavior in leach's storm-petrels as a defense
against slaty-backed gulls. The Auk 103(1):14-22.
Watson MJ, Wilson DR and Mennill DJ (2016) Anthropogenic light is associated with
increased vocal activity by nocturnally migrating birds. The Condor 118(2): 338-344.
Watson R (2017) Ringing at Ngulia to map avian migration. Swara April-June 2017: 50-54.
Available at: safring.birdmap.africa/papers/Ringing at Ngulia.pdf
Watts BD, Reed ET and Turrin C (2015) Estimating sustainable mortality limits for shorebirds
using the Western Atlantic Flyway. Wader Study 122(1): 37-53.
West KE, Jablonski MR, Warfield B, et al. (2010) Blue light from light-emitting diodes elicits a
dose-dependent suppression of melatonin in humans. Journal of applied physiology
110(3):619-626.
Natator depressus (Garman, 1858)
found. Northern Territory Naturalist 19:51-53.
Wilson P, Thums M, Pattiaratchi CB, et al. (2018) Artificial light disrupts the nearshore
dispersal of neonate flatback turtles Natator depressus. Marine Ecology Progress Series
600:179-192.
Wilson P, Thums M, Pattiaratchi CB, et al. (2019) High predation of marine turtle hatchlings
near a coastal jetty. Biological Conservation 236(2019):571-579.
Wiltschko W and Wiltschko R (1999) The effect of yellow and blue light on magnetic
compass orientation in European robins, Erithacus rubecula. Journal of Comparative
Physiology A 184:295-299.
Winger BM, Weeks BC, Farnsworth A, et al. (2019) Nocturnal flight-calling behaviour
predicts vulnerability to artificial light in migratory birds. Proceedings of the Royal Society
B, 286(1900), p.20190364.
Winter Y, López J and Helversen O (2003) Ultraviolet vision in a bat. Nature 425: 612-614.
Witherington BE (1992) Behavioural response of nesting sea turtles to artificial lighting.
Herpetologica 48:31-39.
Witherington B (1997) The problem of photopollution for sea turtles and other nocturnal
animals. In: Behavioral Approaches to Conservation in the Wild, Clemmons JR and
Buchholz R, Editors. Cambridge University Press: Cambridge, p:303-328.
Witherington BE and Bjorndal KA (1991) Influences of artificial lighting on the seaward
orientation of hatchling loggerhead turtles Caretta caretta. Biological Conservation
55(2):139-149.
Witherington B and Martin RE (2003) Understanding, Assessing, and Resolving Light-
Pollution Problems on Sea Turtle Nesting Beaches Florida Fish and Wildlife Conservation
Commission FMRI Technical Report TR-2: Jensen Beach, Florida. 84.

UNEP/CMS/COP14/Doc.30.4.4/Annex 3
143
Yoh N, Kingston T, McArthur E, et al. (2022) A machine learning framework to classify
Southeast Asian echolocating bats. Ecological Indicators 136: 108696.
Yong DL, Heim W, Chowdhury SU, et al., (2021) The State of Migratory Landbirds in the
East Asian Flyway: Distributions, Threats, and Conservation Needs. Frontiers in Ecology
and Evolution 9:613172.
Zhao X, Zhang M, Che X, et al. (2020) Blue light attracts nocturnally migrating birds. The
Condor 122: 1-12.
Zielinska-Dabkowska K and Xavia K (2019) Global approaches to reduce light pollution from
media architecture and non-static, self-luminous LED displays for mixed-use urban
developments. Sustainability 11: 3446.
